We have already seen threats to the Clean Air Act of 1970, the Clean Water Act of 1972 from the Trump Administration. Now the Endangered Species Act of 1973 (16 U.S.C. 1531-1544) faces severe risks.
The Washington Post has reported that the Trump Administration is saying that scientists’ ability to revive extinct species through biotechnology justifies relaxing legal protections. The Post quoted Interior Secretary Doug Burgum as saying that innovation – not government regulation — will save species. He has already met with Dallas-based Colossal Biosciences about using its animals in federal conservation efforts, as well as for potential species restoration. I note that having a few “engineered” specimens living in a zoo is not the goal of the Act or sensible biodiversity conservation programs.
This is just the Administration’s latest maneuver aimed at reducing the Act’s protections, which have been in place since adoption of the Endangered Species Act in 1973. The Fish and Wildlife Service — an agency in the Interior Department — has sought White House comments on a proposed redefinition of “harm” under the act. The term is not defined in the text of the Act, so a rule change could allow for significant reductions in protections, especially regarding listed species’ habitats.
Already, President Trump and his administration have overridden endangered species protections. First, he demanded that the Bureau of Reclamation open water transfer systems to drain water from a Northern California river system to southern California. Ostensibly the action was to protect the area from the devastating wildfires, although scientists declared that a lack of water for firefighters was not the reason the fires caused so much damage. The water had been stored, in part, to protect the habitat of the delta smelt.
President Trump also has revived the long-dormant “God Squad.” a federal committee that can override protections for endangered species. Members include Secretary Burgum and five other high-level officials. It was created by Congressional amendment in the late 1970s, during the fight over whether to build the Tellico Dam on the Tennessee River. It is empowered to approve projects even if they result in the extinction of a species.
In February, Interior Secretary Burgum also rescinded guidance adopted by the Biden Administration aimed at minimizing ship strikes on the Rice’s whale, one of the most endangered marine mammals. He has also ordered staff to consider economic factors when deciding habitat protections.
Other threats came earlier. Elon Musk’s SpaceX launch site is only about 10 miles from Aransas National Wildlife Refuge, which provides winter habitat for one of the “iconic” endangered species, whooping cranes. The Midwestern population of piping plovers is also listed as endangered; it winters along the Gulf coast, including at Aransas. The Refuge is home to 400 bird species, primarily ducks, herons, egrets, ibises, and roseate spoonbills. The few studies of noise impacts on birds focus on nesting – which neither whoopers nor plovers engage in while at Aransas … Still ….
Another refuge — in the middle of the Pacific Ocean – is also under threat from rocket activities. The Post reports that the U.S. Space Force – a branch of the U.S. Air Force – will soon publish an Environmental Assessment regarding plans to build two landing pads on Johnston Atoll. The facilities are intended to expedite movement of military cargo around the globe – by transporting it on large commercial rockets. Johnson Atoll is an unincorporated U.S. territory consisting of four tiny islands about 800 miles southwest of Honolulu. Although tens of thousands of red-tailed tropicbirds, red-footed boobies and sooty terns nest on the atoll, the Space Force said in its notice of intent that it expects the construction and operation of the demonstration project will have no significant environmental impact. This finding has been criticized by several organizations, including the Conservation Council for Hawaii, National Wildlife Refuge Association, and Union of Concerned Scientists. See also this statement by the American Bird Conservancy.
red-footed booby adult & nestling on Johnson Atoll; photo by Jordan Akiyama, USFWS via Flickr
One concern is that construction and operation could re-introduce various invasive species. The Post mentions yellow crazy ants; their acids can cause deformities in birds and, in some cases, deadly infections. The U.S. Fish and Wildlife Service spent a decade eradicating the ants. I note that rats very often are introduced to remote islands by cargo ships and are a significant threat to ground-nesting birds.
red-tailed tropic bird swarmed by yellow crazy ants – on Johnson Atoll; photo by Sheldon Plentovich USFWS via Flickr
Congressional Republicans – who now control both houses of the legislature — are preparing amendments to the Endangered Species Act that would slash protections for at-risk species that are – or might later be – qualified for listing under the Act. One approach is to legislatively remove, or “delist,” those species that have gotten in the way of various activities. The Post names gray wolves and grizzly bears, which ranchers say prey on livestock; plus a lizard in Texas oil country; and the northern long-eared bat, which lives in forests that the timber industry wants to log.
range of northern long-eared map in US & Canada
Citing the fact that only 3% of listed species have recovered, Representative Bruce Westerman of Arkansas, Chairman of the House Committee on Natural Resources, wants to amend the Act to give more power to states. He also plans to limit courts’ power to review agencies’ decisions to remove protections for plants and animals.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
One of the pathways is wooden handicrafts – identified as a pathway more than a decade ago but only partially regulated. The other is new: wood used to decorate aquaria. Two new papers demonstrate that both carry multiple and diverse taxa of fungi. At least 30 have never been detected before in the US. They include both plant and human pathogens.
Both sets of authors call on the U.S. Department of Agriculture to remedy ineffective regulations. However, it appears unlikely that APHIS will be able to do so now, when its budget and staff are being cut. (Lawsuits might restore some of these resources.) Extending APHIS’ authority to regulate organisms that are not plant pests would require Congress to adopt new legislation.
Live Pathogens Imported in Wooden Handicrafts
In 2022, Jason Smith and others (full citation at the end of this blog) published an analysis of the viability and diversity of fungi brought to the U.S. in imported wooden handicrafts. They isolated 47 fungal taxa originating from at least seven countries on three continents. All remained viable despite being subjected to various phytosanitary requirements. Fourteen were plant pathogens; 17 were human pathogens; several were producers of mycotoxins. Three taxa have not been reported before in North America: Bipolaris austrostipae, Paecilomyces formosus, and Xylaria badia. All three are plant pathogens. P. formosus is a human pathogen as well.
Three quarters of the taxa have tolerances that would increase their likelihood of surviving standard heat or fumigation treatments.
Smith et al. point out that wood from sources other than China are subject only to general permit requirements outlined here. Only if pests are detected during port inspections are quarantine actions taken.
APHIS has certified more than a thousand Chinese exporters of handicrafts that incorporate wood, straw, or other biological components. APHIS encourages importers to buy products from these businesses. However, importers may choose other sources. In that case, the product must be treated before entry.
However, as Smith et al. point out, regulation and treatments are focused on arthropods. They do not address the risk from disease pathogens. Smith et al. conclude that these regulations are insufficient to protect plants from damage.
A second issue is that USDA has no authority to regulate organisms that pose a risk to non-plant hosts, including humans. This is especially worrying in this case because many of the handicrafts being intended for food preparation and distribution. Others are handled by purchasers during crafting activities, or used in bath and beauty products.
photo by Sean Welton via Flickr
Live Fungi Imported in Decorative Wood for Aquaria
A second study has expanded the types of material raising concern. The Minnesota Invasive Terrestrial Plant and Pests Center sponsored research that confirms that pieces of wood imported to decorate aquatic and terrestrial aquaria support live fungal-like organisms. The scientists worry that the wood – and the organisms it harbors – might be discarded in a way that facilitates escape and establishment of these organisms. Another possible route of escape is if the water from these mini-habitats is dumped into surface waters.
Blanchette, Rajtar, Lochridge and Held (2025; full citation at end of this blog) obtained 44 samples of such wood from on-line sellers. Some samples had evidence of fungal infestation. Many of the wood pieces were extensively degraded, with large holes, some of which held mud or sand.
The scientists isolated 202 cultures representing 123 fungal taxa in the Ascomycota, Basidiomycota, and Mucoromycota. They detected no Oomycota. The organisms included 30 or 31 species that have not previously been reported in the United States. Twenty-one species are potential plant pathogens, 37 species are wood decay fungi. Twenty-four taxa appear to be previously unknown.
The origins of the wood pieces have not been revealed by the sellers. The scientists believe wood might have come from China, Vietnam, Thailand, and possibly other Asian countries.
Blanchette et al. note that many fungal-like pathogens that have caused devastating diseases in North American forests came from Asia, although not all were introduced directly from there. They name as examples chestnut blight, white pine blister rust, Dutch elm disease, Port-Orford cedar root disease, sudden oak death, and laurel wilt. [Brief descriptions of all these diseases can be found here.] These past introductions occurred via transport of soil, timber, wood products, living trees, or other plant material.
Blanchette et al. cite Smith et al. regarding detection of novel fungal pathogens of both plants and people on imported wooden handicrafts. They cite Brasier, Jung, and others for the likely Asian origins of many Phytophthora species (see citations at the end of this blog). They note the risk associated with the many undescribed species found in that region. In agreement with many others, Blanchette et al. suggest that fungal pathogens pose a very high risk for the U.S. due to rapid emergence of new diseases, low resistance in host populations, and limited surveillance infrastructure for detection.
The Blanchette et al. study was prompted by detection of Xylaria apoda growing on wood submerged in aquariums located in two states which are quite far from each other – Minnesota and Colorado. Despite the pieces of wood having been dried, shipped and stored for a long period during the import process, the fungus remained viable and was producing fruiting bodies. In total, they isolated eight species as known pathogens of agricultural crops and trees. They also report other fungi that might have potential to be plant or human pathogens. Blanchette et al. express specific concerns about possible impacts of the saprophytic taxa on ecosystem functions. That is, wood decay communities could be adversely affected by changes to biomass degradation and native wood-inhabiting insects.
Blanchette et al. point out that their detections came from 44 samples, which represent a very small fraction of the wood being imported for these purposes. Nevertheless the researchers detected impressive quantities and diversity of viable fungi.
Although they did not isolate any Oomycota species, Blanchette et al. say the presence of mud and soil indicates this type of wood could be a pathway for introduction of various exotic Phytophthora – which reside in aquatic and wet soil environments. They call for additional sampling and investigation using more selective methods of isolating Phytophthora species to determine if this could be a successful avenue for importing species of plant pathogenic Oomycota.
Blanchette et al. recommend that people who purchase wood for aquaria repeatedly soak and rinse the wood in water before putting it into an aquarium. This helps eliminate some of the heartwood extractives from the tropical woods and reduces water discoloration in aquariums, as well as possible toxicity to fish and plants. They warn that disposal of the water in contact with this wood into waterways or outdoors could easily release fungal species or Phytophthora spp. that might be in the wood.
Blanchette et al. say their results support earlier indications that current regulations to prevent the importation of non-native fungi on decorative woods used in aquariums are ineffective. In this investigation alone, they cultured more than 100 different live taxa that survived any fumigation or sterilization treatment. They note that scientists have repeatedly called for stronger phytosanitary regulations on imported wood.
One important step they suggest is increasing biosurveillance at the global level. They also suggest prohibiting importation of fungi and fungal-like organisms via this pathway before they become serious problems in their new environment. I concur with these suggestions – with the caveat that while the importation ban is in effect, APHIS and other agencies with authority over invasive species threats to non-plant resources should assess the risks and identify what steps each should take to address them.
[For the history of earlier critiques of weak regulation of imported wood, see blogs on this site under the category “wood packaging” and Fading Forest reports Two and Three (links at the end of this blog). For my critique of regulation of pathogens, see here or contact me.
SOURCES
Blanchette, R.A., Rajtar, N.N., Lochridge, A.G. et al. 2025. Intercontinental movement of exotic fungi on decorative wood used in aquatic and terrestrial aquariums. Scientific Reports 15, 9142. https://doi.org/10.1038/s41598-025-94540-x
Smith, J.A., T. Quesada, G. Alake, N. Anger. 2022. Transcontinental Dispersal of Nonendemic Fungal Pathogens through Wooden Handicraft Imports. mBio July/August 2022 Volume 13 Issue 4 10.1128/mbio.01075-22
Background sources
Brasier, C. M. 2008. The biosecurity threat to the UK & global environment from international trade in plants. Plant Pathol. 57, 792–808.
Brasier, C. M., Vettraino, A. M., Chang, T. T. & Vannini, A. 2010. Phytophthora lateralis discovered in an old growth Chamaecyparis forest in Taiwan. Plant. Pathol. 59, 595–603.
Jung, T., B. Scanu, C.M. Brasier, J. Webber, et al. 2020. A survey in natural forest ecosystems of Vietnam reveals high diversity of both new & described Phytophthora taxa including P. ramorum. Forests 11, 93.
Jung, T., Horta Jung, M.; Webber, J.F. et al. 2021. The destructive tree pathogen Phytophthora ramorum originates from the Laurosilva forests of East Asia. J. Fungi 7, 226.
Jung, T., Milenković I, Balci Y, et al. 2024 Worldwide forest surveys reveal forty-three new spp in Phytophthora major clade 2 with fundamental implications for the evolution & biogeography of the genus & global plant biosecurity. Stud. Mycol. 107, 251-388.
Roy, B. A. et al. 2014. Increasing forest loss worldwide from IAS pests requires new trade regulations. Front. Ecol. Environ. 12, 457–465.
Wingfield, M. J., Brockerhoff, E. G., Wingfield, B. D. & Slippers, B. 2015. Planted forest health: The need for a global strategy. Science 349, 832–836.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
Erythrina caffra one of the native tree species in South Africa killed by PSHB. photo by Coana/Riti via Flickr
Introductions of bark and ambrosia beetles (Coleoptera: Curculionidae, Scolytinae) have significantly increased over the past century. Surveys conducted at borders and ports of entry around the world have shown the majority of beetles intercepted were scolytines. These insects are highly destructive on their own. Also, they can carry pathogenic fungal symbionts that can have devastating effects on the trees they attack.
One or more species in a complex in the Euwallacea genus have become established in countries around the world. One of these, the polyphagous shot hole borer (Euwallacea fornicatus; PSHB) and its associated fungus (renamed from Fusarium euwallaceae to Neocosmospora euwallaceae) is threatening havoc in South Africa about a decade after its establishment (Townsend, Hill, Hurley, and Roets. 2025).
Over this brief period PSHB/Fusarium disease has spread from two introduction sites – Pietermaritzburg, in KwaZulu-Natal Province, and Cape Town, in Western Cape Province – to all but one of the country’s nine provinces. It has become established in four of five forest types studied – Afrotemperate, coastal, sand, and swamp forests. It has not established in mangrove forests. (The Western Cape Province is home to its own “floral kingdom”. The kingdom’s charactersitic fynbos flora is a heathland habitat, not a forest one.)
Townsend and colleagues established a network of 78 monitoring plots in the Western Cape and KwaZulu-Natal provinces. The sites reflected a variety of natural and human impacts.
tree infested by PSHB/Fusarium disease in KwaZulu-Natal Botanical Garden, Pietermaritzburg. Photo from website of Greenpop.org
By monitoring these plots over five years (2019 – 2024), Townsend and colleagues have demonstrated that the beetle/fungus complex and resulting “Fusarium disease” is spreading and intensifying. The number of infected trees rose from 100 to 176 over the five years – a mean increase of 0.6% per year. The number of PSHB entry holes increased by over 10% annually. The number of plots containing infected trees roughly doubled from 23 in 2019 (29% of the 78 plots) to 48 (60%) in 2023.
By the end of the study, 29% of the 148 species sampled had been infected. This represented 43 species and 7 unidentified trees infected. Trees of eight native species died, , although one — Diospyros glabra (Ebenaceae) – resprouted after the main bole died.
In addition to the eight species known to suffer mortality, another 18 species were found to be able to support PSHB reproduction. Townsend and colleagues worry that, as the infestation spreads and intensifies, some of these species might also succumb. They mention specifically Erythrina caffra (coral tree), which is prevalent in coastal forest ecosystems across South Africa.
Most of the hosts are in the same families as those identified earlier by Lynch et al. (2021), e.g., Ebenaceae, Fagaceae, Fabaceae, Malvaceae, Podocarpaceae, Rutaceae, Sapindaceae and Stilbaceae.
Disease progress, speed of death, and visibility of symptoms varied not only between species, but sometimes among individuals of the same species. Some trees died rapidly. Townsend and colleagues say it is impossible to predict which individuals will succumb to infection.
There is, though, a clear frequency-dependent relationship between trees and beetles. Sites with higher relative abundance of host trees also had a higher proportion of infected trees, on average. The number of PSHB holes per species and per plot both increased to a larger extent at these same sites.
Individual trees’ traits influenced the severity of infestations (measured by the number of PSHB entry holes). Larger trees, those with a less healthy canopy, and those farther from a water source suffered more attacks. (This last finding differs from others’; Townsend et al. speculate that in the absence of flood-stressed trees, drought-stressed trees might be more attractive to ambrosia beetles.)
native tree in Tsitsikama National Park; photo by F.T. Campbell
Characteristics of the monitoring plots also affected disease progression. Higher proportions of trees became infected when they grew in plots that were closer to source populations, or that contained a higher proportion of host species as distinct from non-host species. The proportion of trees infected decreased in plots with higher tree densities or tree species richness.
As of 2023, “Fusarium disease” is more widespread and intense in KwaZulu-Natal than in the Western Cape. In KwaZulu-Natal 0.11% of monitored trees are infected compared to 0.06% in the Western Cape. The number of infected trees rose twice as fast over the five years in KwaZulu-Natal – ~6%, than in Western Cape – 3%. While all KwaZulu-Natal plots contained infected trees, three of 11 monitoring sites in the Western Cape did not. Townsend and colleagues believe that the most likely explanation is that PSHB arrived in KwaZulu-Natal earlier (as far back as 2012 as opposed to 2017 in Western Cape). Another possible factor is that source populations of infected trees are indigenous trees within the forest in KwaZulu-Natal whereas, in the Western Cape, they are often non-native trees planted in urban areas far from the study plots. Also, forests in KwaZulu-Natal are fragmented while, in Western Cape, the study forests are nearly contiguous. Townsend et al. conclude that the disease will spread and intensify in Western Cape as additional source populations become established in the forest.
locations of PHSB/Fusarium disease in Cape Town, South Africa – West of the study sites; map from City of Cape Town
As of 2023, the proportion of trees infected appears to be small — 7.6% of the 2,313 trees monitored. Only 11 trees in the monitored plots have died. However, the longer PSHB is active in the environment the more trees it will infest, the higher its impact will be on hosts, and the higher the number of dispersing individuals produced. This will substantially increase the chances and rates of additional areas becoming infected, especially in areas close to infestations – e.g., cities. They fear that in the future impacts will increase as progressively more competent host individuals are infected. Therefore, they emphasize the importance of mitigating PSHB increase in natural ecosystems, even in already infected areas.
Townsend and colleagues urge phytosanitary officials and resource managers to prioritize surveillance and management on the families containing several host species (above) and within plant communities in which they predominate. Managers must also be alert to new reproductive hosts for the beetle that appear as the infestation spreads and intensifies.
The situation could be worse than described; the Townsend et al. study did not examine how the invasion might affect eco-regions outside these two provinces. Because the PSHB has such a broad host range, hosts can die quickly, and South Africa provides ideal climatic conditions, this bioinvader could cause severe ecological effects on most indigenous forest types as well as agriculture and urban trees throughout Africa.
SOURCES
Lynch, S.C., A. Escalen, and G.S. Gilbert. 2021. Host evolutionary relationships explain tree mortality caused by a generalist pest-pathogen complex. Evol Appl 14:1083 – 1094. https://doi.org/10.1111/eva.13182
Townsend, G., M. Hill, B.P. Hurley, and F. Roets 2025. Escalating threat: increasing impact of the polyphagous shot hole borer beetle, Euwallacea fornicatus, in nearly all major South African forest types. Biol Invasions (2025) 27:88 https://doi.org/10.1007/s10530-025-03551-2
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
results of invasion by emerald ash borer (photo courtesy of Nathan Siegert, USFS ); one of the woodboring beetles found to be so damaging to hardwood trees
Over the last nine years scientists have made significant progress in identifying aspects of insect-plant host relationships that play important roles in determining how much damage an introduced, non-native pest is likely to cause within forest ecosystems in the United States. Predicting which introductions will probably cause the greatest damage is vitally important because scientists, phytosanitary officials, and resource managers cannot address all the hundreds of established insects, much less the thousands which might be introduced. This shortfall increases with each surge in import volumes (see my previous blogs about wood packaging by scrolling down the website below the “Archives” to “Categories”, then find “SWPM”), proliferation of goods types and source areas, and cutbacks in funding.
I hope USDA APHIS and Forest Service are adjusting their procedures to apply the scientists’ path-breaking findings.
Their progress will help protect our forests. I apologize if I seem ungrateful — but we need similar progress in managing plant pathogens. Consider the damage caused by chestnut blight, white pine blister rust, Dutch elm disease, sudden oak death, beech leaf disease … (All these and other pathogens are described briefly here.) Understanding the universe of introduced fungi, water molds, nematodes, viruses, etc., is per se much more challenging. Ashley Schulz points out that among the complications are pathogens’ complex life cycles, and possible new relationships with vectors.
Undertaking this analysis will be set back decades if agencies’ resources – funds and staffs – are decimated during the current “downsizing” of government. We must speak up!!
At least regarding non-native insects that attack North American tree species, scientists’ analyses promise a new ability to set priorities. This should improve the efficacy of phytosanitary programs – if government downsizing is not allowed to destroy USDA’s scientific, regulatory, and resource management programs.
We must speak up!!
What science tells us now
Schulz et al. (2025) summarize current findings. (Full citation to all references appear at the end of the blog.)
Earlier, scientists sought to find commonalities associated with introduced insects that caused high impacts on North American conifer trees [Mech et al. (2019)] and hardwoods (angiosperms) [Schulz et al. (2021)] (Full citations at the end of the blog; earlier blogs posted here and here.) Both studies found that the time elapsed since tree species in North America diverged from the host plants of the insects in their native range (i.e., host evolutionary history) is a diagnostic factor. This factor best predicted non-native insect impact compared to the other factors that were significant for conifer and hardwood specialists. For conifers, the other significant factors included the shade and drought tolerance of the North American host plants and whether there was a related insect native to North America on the same hosts that the non-native insect impacted. For hardwoods, another important factor explaining a specialist insect’s impact is if the insect is a wood borer, especially a scolytine beetle. The wood density of the North American host plant was also considered a significant factor when predicting impact of the non-native insect.
In 2022, Uden et al. applied the divergence time method to insect species not yet introduced to North America that might attack conifer species. They hoped to identify both insects posing the greatest hazard and tree species most vulnerable to introduced pests.
Now, a new team again led by Ashley Schulz and Angela Mech (see Schulz et al. 2025) has applied a similar approach to a more comprehensive range of pest-host relationships, including the pests that specialize on host plants and pests that feed on a broader array of hosts. Some feed on both conifers and hardwoods. They found that:
It is possible to quantify insect host breadth and identify the cutoff where “specialists” and “generalists” diverge. Specifically, the split occurs around 2,250 cumulative million years, where insects that feed on hosts that add up to less than that have narrow host breadth (i.e., “specialists”) and insects that feed on hosts that add up to more than that have broad host breadth (i.e., “generalists”). This technique also helps categorize insects that fall within the middle range of host breadth and are traditionally difficult to classify as either specialists or generalists based on differing qualitative definitions of the terms.
Insects that use more hosts in their native range also tend to use more hosts in the introduced range (North America). However, many of these insects utilized fewer hosts in the introduced range compared to the native range. This shrinkage was not universal, however; about 30% of insects increased their host breadth in the introduced range. Most of these fed on a single species in their native range but attacked additional species in the same family in North America. The corresponding i-Tree Pest Predictor tool uses the list of hosts in the insect’s native range and these models to determine the insect’s likelihood that it would cause high impact, as well as each North American tree species’ susceptibility to the insect entered into the tool.
Certain feeding guilds had – on average — a significantly narrower host breadth in North America than in their native ranges. These were gall makers (13 species analyzed); sap feeders (120 species); and wood borers (35 species). In contrast, host ranges did not differ for folivores (68 species), reproductive feeders (7 species), and root feeders (5 species). Still, we know that wood borers, as a group, have caused enormous damage to a range of North American tree taxa (see emerald ash borer, redbay ambrosia beetle, invasive shot hole borers (all described briefly here). Again, the i-Tree Pest Predictor tool can help identify the threat to particular tree species.
Of course, APHIS should not disregard pests with narrow host ranges; several have caused enormous damage.
red spruce (Pikea rubens) in Great Smoky Mountains National Park; photo by famartin via Wikipedia. Red spruce is the species found to be most vulnerable by the Uden et al. study
Schulz et al. (2025) developed models for three groups of introduced herbivorous insects that feed on trees:
1) conifer specialists (based on analysis of 69 species);
2) hardwood specialists (based on analysis of 141 species);
3) hardwood generalists (based on analysis of 30 species).
Because of their quantification of host breadth, they defined the “specialist” group more broadly than is commonly done, e.g., an insect that feeds on the three families Betulaceae, Fagaceae, and Juglandaceae would be considered “specialists” because all three host families are in the Fagales clade.
Tree relatedness was the only significant explanatory factor for all three host breadth categories. As determined in the previous studies, North American host tree species that were too closely or distantly related to the insect’s hosts in its native range were less impacted than hosts that diverged somewhere in the middle – the “Goldilocks” range. The divergence period differs among the three pest-risk categories: 3–4 million years ago for conifer specialists, 5–9 million years ago for hardwood specialists, ~1–2 million years ago for hardwood generalists. Schulz et al. suggest that the reason why the peak probability of high impact differs among these groups is that different feeding guilds cause the most damage to the specific host category, and each feeding guild is challenged by different tree host defenses. Bark and wood boring beetles (the hardwood specialists with the greatest impact) must overcome lethal constitutive and induced tree defenses in order to survive for long periods in the cambial layer. These insects have adapted the ability to locate and select poorly defended individuals in the host population. Folivores (i.e., the generalists with the highest impact) adapt to plant chemistry and trichomes (hair-like or scale-like outgrowths), or can avoid host defenses by moving off the foliage. Sap feeders (which include many high impact conifer specialists) are usually tolerated by trees, unless they stimulate hypersensitive reactions or vector pathogens.
Of course, scientists’ estimates of how long ago tree taxa diverged from common ancestors differ. Fortunately, Uden et al. (2022) found that these differences only rarely affect the predicted impact of a non-native insect – at least in the case of the 62 European insects and 47 North American conifer species they analyzed. In only 1.37% of the 2,914 pairs analyzed did the predicted risk differ depending on which source phylogeny was used. These cases were associated with 27 conifer-specialist insects and 9 conifer hosts. The article does not tell us which pest/host pairs these are but, overall, this paper demonstrates that the estimate differences in the phylogenetic trees does not differ enough to be problematic when forecasting insect impact.
Changes Needed in the Way Agencies Set Priorities
Schulz et al. (2025) urged agencies to stop relying only on insect traits as the basis for developing models & phytosanitary regulations. The only insect trait that predicted impact is the insect’s feeding guild. Considering hardwoods, they found that wood borers pose the greatest risk among specialists to hardwoods; folivores among generalists. While sap feeders do not cause statistically higher damage on hardwood tree species, four of the seven high-impact conifer specialists are sap feeders (hemlock woolly adelgid, balsam woolly adelgid, red pine scale, and spruce aphid). Therefore, the i-Tree Pest Predictor tool incorporates consideration of whether a pest of conifers is a sap feeder.
Schulz et al. (2025) also caution agencies against relying on just the number of hosts an insect might exploit. Assessors must consider the range of underlying plant chemistry / host defenses that the insect encounters. They found that hosts that are shade tolerant are more susceptible to high impact from conifer specialists and hosts that have intermediate to no shade tolerance are more susceptible to high impact from generalists.
Uden et al. (2022) identified a possible weakness in USDA efforts to prioritize pest prevention targets. They found that APHIS’ Prioritized Offshore Pest List included only 12 conifer specialists from Europe among the 150 species listed. They go on to note that while sap feeders constitute 53% of tree pest species established in the U.S., APHIS listed none. The models applied by Uden, Schulz, and Mech do not consider whether the insect is likely to become established. Improving our understanding of the many factors influencing an insect’s likelihood of being transported to North America or becoming established requires additional research. This might eventually lead to a usable tool for predicting this aspect of bioinvasion by forest pests.
There is an urgent need for such a tool. As Uden et al. noted, they found that 66% of the insect species they analyzed fell into the “high impact” category. This is a much higher proportion than estimates based on earlier studies, so identifying which of these insects are likely to establish versus not establish in North America can provide more resolution and help identify which insects are going to be most problematic.
Mature Fraser fir killed by balsam woolly adelgid; Clingman’s Dome; photo by Ben Ramsey via Flickr
Tree species at risk
The analysis undertaken by Uden et al. determined that three conifer species face a high level of hazard from European insects if they are introduced. They identified particularly high threats to two species, Fraser fir (Abies fraseri) and Carolina hemlock (Tsuga caroliniana). The fir is determined to be vulnerable to 17 insect species which are predicted to have high likelihood of a high impact. The hemlock is highly vulnerable to one of the insect species they sampled. They note that both of these conifers have a limited geographic range and ecological habitat, so they likely have a relatively narrow genetic pool. A third species said to be at elevated risk is red spruce (Picea rubens) – which, although more widespread, is also under attack by a non-native insect. All three species fit earlier finding by Mech et al. that conifer trees with high shade tolerance but low drought tolerance more vulnerable to non-native pests. In none of these cases do Uden et al. mention that the tree species have already been severely diminished by established non-native insects – i.e., balsam woolly adelgid on the fir (above), hemlock woolly adelgid on the Carolina hemlock. The Schulz/Mech team is working to refine methods for identifying tree species and regions at greatest risk.
Meanwhile, Uden et al. have suggested that phytosanitary authorities and forest managers apply their findings to identify the European herbivorous insects that pose the greatest threat to North American conifer species. They should identify Palearctic tree species that fall within the high-impact “Goldilocks” zone of divergence times in relation to specific North American tree species, then identify the insects that feed on those Palearctic trees. These insects would presumably pose the highest predicted hazard to those North American tree species. They suggest that species so identified should be added to the USFS’ list of species targetted by its wood borer early detection program. To address likelihood of introduction, they suggest incorporating data on insect species commonly intercepted at ports – an indication of high propagule pressure. There will always be exceptions though. For example, Ips typographus feeds on spruce and has been frequently detected at the ports, but it has not established in North America.
For those focused on identifying species or ecoregions at greatest risk, Uden et al. suggest scientists use several sources to identify vulnerable vegetation communities. Sources suggested include USFS Forest Inventory and Analysis (FIA) and NatureServe Explorer plant community descriptions) that have relatively high-value tree species predicted to be at risk from introduced species.
SOURCES
Aukema, J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, and S.J. Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the Continental United States. BioScience 60(11): 886-897. https://doi.org/10.1525/bio.2010.60.11.5
Mech, A.M., K.A. Thomas, T.D. Marsico, D.A. Herms, C.R. Allen, M.P. Ayres, K.J. K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, & P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecology and Evolution 9(21): 12216–12230. https://doi.org/10.1002/ece3.5709
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions 23: 3921-3936. https://doi.org/10.1007/s10530-021-02621-5
Schulz, A.N., N.P. Havill, T.D. Marsico, M.P. Ayres, K.J.K. Gandhi, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, K.A. Thomas, P.C. Tobin, D.R. Uden, A.M. Mech. 2025. What Is a Specialist? Quantifying Host Breadth Enables Impact Prediction for Invasive Herbivores
Uden, D.R., A.M. Mech, N.P. Havill, A.N. Schulz, M.P. Ayres, D.A. Herms, A.M. Hoover, K.J.K. Gandhi, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, K.A. Thomas, P.C. Tobin, C.R. Allen. 2022. Phylogenetic risk assessment is robust for forecasting the impact of European insects on North American conifers. Ecological Applications 33(2): e2761. https://doi.org/10.1002/eap.2761
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
Scientists in New Zealand are saying explicitly that a forest’s unique mixture of species matters when considering the future. This mixture is the result of the forest’s evolutionary history. Losing members of the biological community reduces the forest’s ability to respond to current and future stresses – its resilience.
New Zealand’s forests are part of the broader legacy of the ancient supercontinent of Gondwanaland – the island nation’s plants have close relatives in South America, the Pacific Ocean islands, and Australia. Still, these forests are unique: 80% of New Zealand’s plant species are endemic. The forests are also species-rich. The warm temperate evergreen rain forests of the North Island are home to at least 66 woody plant species that can reach that reach heights above six meters (Simpkins et al. 2024).
These forests have been severely changed by human activity. In just ~ 750 years people have cut down approximately 80% of the original forest cover! (Simpkins et al. 2024) Of the eight million hectares of surviving native forest, a little over five million hectares is managed for the conservation of biodiversity, heritage, and recreation. Another 2 million hectares are plantations of non-native species.
sites in New Zealand where pine plantations are “wilding”
All these forests are challenged by introduced mammals – from European deer to Australian possums. Climate change is expected to cause further disturbance, both directly (through e.g., drought, extreme weather) and indirectly (e.g., by facilitating weed invasion and shifting fire regimes) (Simpkins et al. 2024).
Pathogen threats are also common threats to the native trees of the Pacific’s biologically unique island systems. For example, Ceratocystis lukuohia and C. huliohia (rapid ‘ōhi‘a death, or ROD). The latter is killing ‘ōhi‘a (Metrosideros polymorpha) on the Hawaiian Islands. More than 40% of native plant species in Western Australia are susceptible to Phytophthora cinnamomi. Here I focus on two pathogens, kauri dieback and myrtle rust, now ravaging New Zealand’s native flora. No landscape-level treatment is available for either pathogen.
When considering this suite of challenges, Simpkins et al. focus on these two pathogens’ probable impact on forest carbon sequestration. They worry in particular about erosion of the forests’ resilience due to loss of “ecological memory” – the life-history traits of the species (e.g., soil seed banks) and the structures left behind after individual disturbances.
one of the largest remaining kauri trees, “Tane Mahuta”, in Waipoua Kauri Forest; photo by F.T. Campbell
Kauri Dieback
The causal agent of Kauri dieback, Phytophthora agathidicida, is a soil-borne pathogen that spreads slowly in the absence of animal or human vectors. The disease affects a single species, Agathis australis (kauri, Araucariaceae). However, kauri is a long-lived, large tree that is a significant carbon sink. It probably modifies local soil conditions, nutrient and water cycles, and associated vegetation. Also, kauri has immense cultural significance.
Simpkins et al. note that kauri dieback threatens stand-level loss of A. australis – that is, local extinctions. In the absence of disturbance Kauri trees can grow to awe-inspiring size. In the 19th Century, before widespread logging, some were measured at 20 meters or more in circumference. Consequently, kauri dieback might cause a decline in aboveground live carbon storage of up to 55%. This loss would occur over a period of hundreds of years, not immediately.
Huge kauri are not likely to be replaced by other long-lived emergent conifers (based on an analysis of one species, Dacrydium cupressinum). Instead, kauri are probably going to be replaced by late-successional angiosperms. The authors discuss the ecological implications for levels of carbon storage and proportions of trees composed of Myrtaceae – exacerbating damage caused by myrtle rust (see below).
The expectation of Simpkins et al. that kauri will suffer at least local extinctions is based on an assumption that no kauri trees are resistant to the pathogen. Fortunately, this might not be true: different Agathis populations show various levels of tolerance to Agathis dieback. Identification and promotion of some levels of resistance could enable A. australis to retain a diminished presence in the landscape.
However, Lantham, et al. make clear that containing kauri dieback remains “challenging,” despite its discovery nearly 20 years ago (in 2006). Scientists and land managers have little information on the distribution of symptomatic trees, much less of the pathogen itself. This means they don’t know where infection foci are or how fast the disease is spreading.
As is often true, the pathogen is probably present in a stand for years, possibly a decade or more, before symptoms are noticed. This means that the current reliance on public reports of diseased trees, or targetting surveillance on easy-to-access sites (e.g., park entrances and along existing track networks), or at highly impacted areas readily identified through aerial methods, fails to detect early stages of infection. Indeed, it seems probable that P. agathidicida had been present in New Zealand’s ecosystems for decades before its formal identification.
The Waipoua forest is one of the largest areas of forest with old kauri stands in the country. A new analysis of aerial surveys done between 1950 and 2019, shows how the forest is changing. The number of dead trees increased more than four-fold and the number of unhealthy-looking trees increased 16-fold over these 70 years. Kauri dieback is now widespread in this forest, especially in areas near human activities like clearing for pasture or planting commercial pine plantations).
Lantham et al. have developed a model which they believe will help identify areas of higher risk so as to prioritize surveillance and inform responses. These could delimit the disease front and help implement quarantines or other measures aimed at limiting the spread of P. agathidicida to uninfected neighboring sites.
I hope New Zealand devotes sufficient resources to expand surveillance and management to levels commensurate with the threat to this ecologically and culturally important tree species.
Leptospermum scoparia; photo by Brian Gatwicke via Flickr
Myrtle Rust
Myrtle rust is a wind-borne disease that affecting numerous species in the Myrtaceae, including some of the dominant early successional species (e.g., Leptospermum spp.). Simpkins et al. expect that myrtle rust might hasten the decline of two such tree species (L. scoparium and Kunzea ericoides). However, these trees’ small size and rapid replacement by other species during succession minimizes the effect of their demise on carbon storage.
Because I am concerned about the irreplaceable loss to biodiversity, I note that Simpkins et al. also feared immediate threats to some trees in the host Myrtaceae family, specifically highly susceptible species such as Leptospermumbullata.
As I reported in a recent blog, a second group of scientists (McCarthy et al.) explored the threat from myrtle rust more broadly. Austropuccinia psidii has spread through Myrtaceae-dominated forests of the Pacific islands for about 20 years.
Trees in the vulnerable plant family, Myrtaceae, are second in importance (based on density and cover) in New Zealand’s forests. Successional shrub communities dominated by the two species named above, Kunzea ericoides and Leptospermum scoparium, are widespread in the northern and central regions of the North Island and in northeastern and interior parts of the South Island. These regions’ vulnerability is exacerbated by the area’s climate, which is highly suitable for A. psidii infection (Simpkins et al. 2024).
McCarthy et al. concluded that ifLeptospermumscoparium and Kunzea ericoides prove to be vulnerable to myrtle rust, their loss would cause considerable change in stand-level functional composition across these large areas. They probably would be replaced by non-native shrubs, which are already common on the islands. Any resulting forest will differ from that formed via Leptospermeae succession.
These authors also worry that the risk to native ecosystems would increase if more virulent strains of the myrtle rust pathogen were introduced or evolved. They note that A. psidii is known to have many strains and that these strains attack different host species.
SOURCES
Latham, M.C., A. Lustig, N.M. Williams, A. McDonald, T. Patuawa, J. Chetham, S. Johnson, A. Carrington, W. Wood, and D.P. Anderson. 2025. Design of risk-based surveillance to demonstrate absence of Phytophthora agathidicida in New Zealand kauri forests. Biol. Invasions (2025) 27, no. 26
McCarthy, J.K., S.J. Richardson, I. Jo, S.K. Wiser, T.A. Easdale, J.D. Shepherd, P.J. Bellingham. 2024. A Functional Assessment of Community Vulnerability to the Loss of Myrtaceae from Myrtle Rust. Diversity and Distributions, https://doi.org/10.1111/ddi.13928
Simpkins, C.E., P.J. Bellingham, K. Reihana, J.M.R. Brock, G.L.W. Perry. 2024. Evaluating the effects of two newly emerging plant pathogens on North Aotearoa-New Zealand forests using an individual-based model. Ecological Modelling, www.elsevier.com/locate/ecolmodel
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
riparian ash killed by EAB; in this case, Mattawoman Creek in Maryland. Photo by Leslie A. Brice
Good news at the recent 33rd USDA Research Forum on Invasive Species. Scientists presented the first study that demonstrates significantly lower ash tree mortality in sites with high parasitism rates of two larval parasitoids, Tetrastichus planipennisi and Spathius galinae.
Their study area is the ash-dominated riparian area along the Connecticut River that flows north to south across the middle of Massachusetts. Knowing in advance that the emerald ash borer (Agrilus planipennis; EAB) would invade the area, scientists established monitoring plot that consisted of marked individual ash trees. EAB was first detected in the southern reach of the riparian area in 2015. It gradually moved north. By 2020 isolated mortality was observed at all sites. Meantime, they released three biocontrol agents – T. planipennis,S. galinae, and Oobius agrilii – early in the invasion at three of the six monitoring sites. These released occurred in 2018 – 2020 and again in 2022.
In 2021 and 2025, the scientists counted the numbers of biocontrol agents in the marked trees or sentinel logs. Thus the first evaluation occurred six years after EAB arrived, three years after the first releases of biocontrol agents.
They found that at southern Massachusetts sites, where EAB density was higher at the time of the biocontrol agents’ initial release, remaining ash grew more slowly than in the North. They believe the trees’ growth rate was suppressed by the trees having fewer resources. They also observed dieback. Smaller trees grew faster, perhaps responding to opening of the canopy as mature ash succumbed to EAB invasion.
The most important finding was that ash mortality at all sites was ~50% or less … not the 90% expected based on experience in the upper Midwest where the EAB invasion occurred before biocontrol agents were developed.
SOURCE
Ash survival and growth response to emerald ash borer invasion in Massachusetts riparian forests: impacts of biological control. Mitchell A. Reed, Jian Duan, Ryan S. Crandall, Roy G. van Driesche, Jeremy C. Anderson, Joseph S. Elkington. Presentation to the 33rd USDA Interagency Research Forum on Invasive Species, Annapolis, Maryland February 25-28, 2025 (The proceedings should be posted online before the end of the year.)
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
discarded pallets next to developed area in Glacier National Park (!); photo by F.T. Campbell
Since July 2015 I have posted nearly 50 blogs about non-native insects introduced via movement of solid wood packaging material (SWPM). Why? Because SWPM is one of two most important pathways by numbers introduced & by impact of the species introduced. (The other pathway is P4P.) To read those earlier blogs, scroll below “archives” to “categories”, choose “wood packaging”.
dead redbay trees in Everglades National Park; killed by laurel wilt vectored by redbay ambrosia beetle
As I have reported in the earlier blogs and in my “Fading Forests” reports (links at the end of this blog), in 2002, the parties to the International Plant Protection Convention (IPPC) adopted an international “standard” to guide countries’ programs intended to reduce the presence of damaging insects in the wood packaging: International Standard for Phytosanitary Measures (ISPM) #15). The U.S. and Canada adopted the standard through a phase-in process culminating in 2006. [For a discussion of the phase-in periods and process, read either of the studies by Haack et al. cited at the end of this blog.] In other words, the U.S. and Canada have implemented ISPM#15 for almost 20 years. China specifically has been subject to requirements that it treat its SWPM even longer – since December, 1998, i.e., more than 25 years.
Unfortunately, ISPM#15 is not intended to prevent pest introductions. As stated in Greenwood et al 2023, “Prior to 2009, the goal of compliance with ISPM 15 was to render the risk of wood-borne pests “practically eliminated,” in 2009 the standard was amended to “significantly reduced”.
Despite almost universal adoption of the standard by countries engaged in international trade, insects have continued to be present in wood packaging. A very high proportion of these infested shipments — 87% – 95% — of the SWPM found by U.S. officials bears the ISPM#15 stamp – that is, is apparently compliant. (See my blogs by clicking on the “Category” “wood packaging” listed below the “Archives”.) The same proportion was found in a narrower study in Europe (Eyre et al. 2018). All the post-2006 examples of infested wood analyzed by Haack et al. (2022) (see below) carry the stamp. I conclude that the ISPM#15 mark has failed in its purpose: to reliably indicate that SWPM accompanying imports has been treated so as to minimize the likelihood that an insect pest will be present.
Dr. Robert Haack, retired USFS entomologist, has twice tried to estimate the “approach rate” of insects in SWPM entering the United States (both studies are cited at the end of this blog). A study published in 2014 that relied on data from 2009 found that U.S. implementation of ISPM#15 was associated with a reduction in the SWPM infestation rate reported of 36–52%. The authors estimated the infestation rate to be 0.1% (1/10th of 1%, or 1 consignment out of a thousand). (See Haack et al. 2014; citation at the end of this blog.)
In their second study, published in 2022, Haack and colleagues found a 61% decrease in rates of borer detection in wood packaging when comparing numbers of wood borer detections in 2003 – before the U.S. implemented ISPM#15 – to those in 2020. Specifically, detections dropped from 0.34% in 2003 to 0.21% in 2020. This decrease occurred despite the volume of U.S. imports rising 68% between 2003 and 2020. (My blogs document a further increase in import volumes over the years since 2020.) In addition, the number of countries from which the SWPM originated more than doubled from 2003–2004 to 2010–2020. This expansion exposes North America to a wider range of insect species that might be introduced, as well as a wider range of individual countries’ effectiveness in enforcing the standard’s requirements (Haack et al. 2022).
These decreases are encouraging. However, Haack et al. (2022) note some caveats:
The reduction in pest presence was greatest during the initial implementation of the program the first phase, 2005-2006 (61%); in subsequent periods pest approach rate inched back up. In the 2010-2020 period, the pest detection rate was only 36% below the pre-ISPM#15 level. Detection rates have been relatively constant since 2005. Does this stasis mean that exporters learned that they could ignore or circumvent the requirements without suffering significant penalties? Or is some of this rise related to increased trade volumes, increasing variety of country of origin for trade, or other global trade patterns unrecognized in the data? (However, see the next bullet point.)
Certain types of commercial goods and exporting countries have consistently fallen short. Specifically, the rate of wood packaging from China that is infested remained relatively steady over the 17 years since 2003. The proportion of consignments with infested wood packaging coming from China was more than five times the proportion of all inspected shipments for this period. In other words, China has had a consistent record of poor compliance with phytosanitary regulations since they were imposed in December 1998. Why is USDA not taking action to correct this problem? (As I note below, DHS CBP has ramped up enforcement efforts.) Some other countries, e.g., Italy and Mexico, have reduced the rate at which wood packaging accompanying their consignments is infested. In fact, Mexico’s improved performance largely explains the overall infestation rate estimate of 0.22% during the period 2010-2010. Mexico’s successes affect the overall statistics in a way that makes other countries’ failure to reduce the presence of pests in wood packaging they ship to the United States far less obvious.
Haack et al. (2022) discuss ten possible explanations for their finding that pest approach rates – as determined by their study — have not decreased more. See the article or my blog about the study.
Although USDA APHIS has not taken steps to strengthen its enforcement, U.S. Customs and Border Protection [an agency in the Department of Homeland Security] has done so twice — see here and here. CBP staff have expressed disappointment that these actions reduced the numbers of shipments in violation of ISPM#15 by only 33% between Fiscal Year 2017 and FY2022. True, more than 60% of these violations consisted of a missing or fraudulent ISPM#15 stamp. However, 194 consignments still harbored live pests prohibited under the standard.
APHIS did agree in 2021 to enable the study by Robert Haack and colleagues, via an interoffice data sharing agreement between USDA APHIS and the Forest Service- this resulted in Haack et al. 2022.
APHIS and CBP also collaborated with an industry initiative to train inspectors that insure other aspects of foreign purchases. The ideas was that CBP or APHIS and their Canadian counterparts would inform importers about which foreign treatment facilities have a record of poor compliance or suspected fraud. The importers could then avoid purchasing SWPM from them. I have heard nothing about this initiative for three years, so I fear it has collapsed.
We lack data on which to base a rigorous analysis
While the two studies by Robert Haack and colleagues are the best available, and they relied on the best data available, the fact is that those available data do not provide a full picture of the risk of pest introduction associated with wood packaging. As pointed out by Leigh Greenwood of The Nature Conservancy in her presentation to 2025 USDA Invasive Species Research Forum, available data have been collected for different purposes than to answer this question. Leigh’s powerpoint is posted here.
Leigh has identified the following data gaps:
In their studies, Haack and colleagues rely on data from the Agriculture Quarantine Inspection Monitoring (AQIM) system. This dataset is based on random sampling of very distinct segments of incoming trade. It is therefore a better measure of insect approach rates than reports of interceptions by either APHIS or CBP.
However, AQIM includes data from only those very distinct segments of trade: perishable goods, SWPM associated with maritime containerized imports, Italian tiles, and “other” goods, AQIM does not contain a segment of trade that includes wood packaging associated with maritime breakbulk or roll-on, roll-off (RORO) cargo. These exclusions have prevented scientists and enforcement officials from determining, inter alia, how great a risk of pest introduction is associated with various types of wood packaging, especially dunnage, as the randomized sample does not include entire pathways for the entrance of dunnage.
Greenwood states that she has not found another country that operates a similar analysis of randomly collected data at ports of entry.
2) USDA does not collect data on consignment size, piece-specific infestation density, nor consignment-wide infestation density. As Haack et al. (2022) point out, reporting detections by consignment doesn’t reveal the number of insects present. If implementation of ISPM#15 resulted in fewer live insects being present in an “infested” consignment, this would reduce the establishment risk because there is lower propagule pressure. However, we cannot know whether this is true.
3) Neither USDA nor CBP reports the inspection effort. Nor do they conduct a “leakage survey” to see how often target pests are missed. This means, inter alia, that we cannot estimate inspectors’ efficiency in detecting infested wood packaging. If their proficiency has improved as a result of improvements in training, inspection techniques, or technology, the apparent impact of ISPM#15 would be under-reported in recent years.
4) USDA does not require port inspectors to report the type of SWPM in which the pest was detected. Leigh participated in an effort that included industry representatives, DHS CBP and USDA APHIS to define the types of wood packaging in legal terminology so that they could be incorporated in the drop-down menu on inspectors’ reporting system. This was first successfully included in the legal glossary within USDA APHIS system of record, ACIR Glossary. Last fall the team was working to integrate the requirement for using these definitions into the inspection data collection system used by DHS CBP, which would then make this data available in Agricultural Risk Management, ARM (see Abstract here for adequate primer on ARM). However, it is unclear now whether the new administration will do so. One potential barrier is that asking the port of entry inspection staff to record these data will add to the time and training required for reporting inspection results.
In summary, Leigh reports that current data systems do not support
estimating probabilities of pest infestation of via volume or type of SWPM (e.g. pallet vs dunnage)
measuring the risk of arrival associated with a specific hazard (in this case, a hazard being a live pest or pathogen associated with SWPM)
extrapolating or supporting findings for some types of wood packaging to other types of wood packaging
Scientists from Canada, Mexico, and the United States have formed a working group under the auspices of the North American Plant Protection Organization (NAPPO). The group is trying to determine whether various types of wood packaging are more likely to harbor pests. This study is currently hampered by the many data gaps, including those Leigh outlined above. The best data available, cited by Haack et al. (2022), found that in maritime containerized shipping, crates were more likely to harbor pests than pallets- however, other forms of SWPM (dunnage, bracing, etc.) had such low sample size that no analysis of those is possible. One of the main objectives of the NAPPO study is to evaluate if dunnage poses the same or higher risk, so this is a major impediment.
Two issues need to be resolved.
One is scientific: why are insects continuing to be detected in wood packaging marked as having been treated? What is the relative importance of insects surviving the treatment versus treatment facilities applying the treatments incorrectly or inadequately?
The second issue is legal and political: what proportion of the detections is due to treatment facilities committing outright fraud – claiming to treat the wood, stamping it with an IPPC stamp, while not actually performing any treatments at all?
Knowing which measures will most effectively solve these quandaries / reduce pest presence in wood packaging depends on knowing what the relative importance of these factors are in causing the problem. The lack of basic data on which to base any analysis certainly hampers efforts to improve protection.
Leigh calls for researchers to recognize these data needs and work to fill them.
•Understand, account for, and communicate data realities
•Work collectively to increase useable data quality
•Use additional research to validate, or to demonstrate disparities
Why Wait for the Science?
In the meantime, however, I assert that more vigorous enforcement efforts by responsible agencies should help reduce the occurrence of fraud. I have suggested the following actions:
U.S. and Canada refuse to accept wood packaging from foreign suppliers that have a record of repeated violations – whatever the apparent cause of the non-compliance. Institute severe penalties to deter foreign suppliers from taking devious steps to escape being associated with their violation record.
APHIS and CBP and their Canadian counterparts follow through on the industry-initiated program described above and here aimed at helping importers avoid using wood packaging from unreliable suppliers in the exporting country.
Encourage a rapid switch to materials that won’t transport wood-borers. Plastic is one such material. While no one wants to encourage production of more plastic, the Earth is drowning under discarded plastic. Some firms are recycling plastic waste into pallets.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
eastern hemlocks in Cook Forest State Forest Pennsylvania; photo by F.T. Campbell
As Eastern hemlock (Tsuga canadensis) suffers high levels of mortality across nearly all its range due to hemlock woolly adelgid (HWA; Adelges tsugae), scientists scramble to determine what the successor forests will look like. The transformation will be stark: from deeply shaded evergreen coniferous forest with a sparse understory to something very different. As this process takes place, most scientists expect cascading effects on not only terrestrial and aquatic wildlife but also onecosystem functions, including soils and nutrient and hydrologic cycles (Dharmadi et al. 2019 Plotkin et al. 2024).
New England
In southern New England, hemlock groves are being replaced by stands of deciduous hardwood forests dominated by black birch (Betula lenta). While birch are expected to continue to dominate, other species comprise at least one third of seedlings in the Harvard Forest experimental sites, primarily eastern white pine (Pinus strobus) and red maple (Acer rubrum). Plotkin et al. (2024) note that conversion of hemlock forests to pine forests would be a less dramatic ecosystem shift since both are evergreen conifers.
symptoms of beech leaf disease; photo by the Ohio State University
In both southern New England and farther north, in Vermont and New Hampshire, maples and American beech have increased in prominence. In the latter case, this is despite the prevalence of beech bark disease and managers’ efforts to suppress beech. I have noted that beech leaf disease now threatens to disrupt this process.
Landowners in the region often seek to get some financial return from their forests before a pest kills the trees. About a quarter of the almost 9,000 ha of hemlock stands in the southern Connecticut River Valley have been harvested as HWA spread into the area. To test the effect of pre-mortality logging of hemlock stands, Plotkin et al. tried to mimic HWA-caused mortality by girdling all the hemlocks in some plots in Harvard Forest. In other plots they harvested most hemlocks and some of the other tree species. The girdled plots had a dramatic increase in standing and downed deadwood and a longer period of elevated understory light levels than the logged plots. They note that standing snags and on-ground dead wood provide critical ecosystem functions. Many wildlife and microbial species depend on dead wood for nutrition and a variety of micro habitats. Plotkin et al. found that the slowly decomposing dead wood also stored a large amount of carbon: girdled plots stored 18% more above-ground carbon than logged sites, even after accounting for carbon stored in harvested wood products.
a beech snag with nesting cavities; photo by F.T. Campbell
The magnitude of these differences might be even larger than demonstrated in this experiment. In New England, hemlocks infested with HWA die over a decade, not the two years seen after girdling. The delayed mortality provides a longer window of opportunity for succeeding vegetation to adapt and preserve higher levels of biodiversity. Plotkin et al. (2024) suggest that logging pest-threatened hemlock forests might remove structural resources that would support forest response to ongoing climate stress and future disturbances.
Considering the disturbed plots’ invasibility by non-native plants, Plotkin et al. (2024) found that more non-native shrubs invaded the girdled plots than the logged plots. They suggest that birds that disperse the shrubs’ fleshy fruits were attracted by perch sites provided by the standing dead trees.
Southern Appalachians
In the Southern Appalachians, post-HWA forests will apparently be quite different. At the USDA Forest Service’ Coweeta Hydrologic Laboratory in the Nantahala Mountain Range of western North Carolina, eastern hemlock died much faster than in New England. Hemlocks comprised more than 40% of the basal area before arrival of HWA (detected in 2003). Within two years all hemlock trees were infested. Half were dead by 2010, 97% by 2014 (Dharmadi et al. 2919).
In some part of the southern Appalachian forests the shrub layer is dominated by Rhododendron maximum (rosebay rhododendron). This dense shrub layer is preventing recruitment of deciduous tree species that had been expected to replace the dead hemlocks. Tree seedlings died rather than grew into saplings. Scientists working in the Coweeta experimental forest attribute the seedlings’ demise to limited access to key resources, e.g., water, nutrients (especially inorganic nitrogen), and light (Dharmadi, Elliott and Miniat 2019).
In the Coweeta Basin, hemlock loss is the most recent of a series of severe disturbances that have apparently led to a cascade of responses in the overstory, midstory, and soil that have promoted expansion of rhododendron. (The earlier disturbances were widespread logging in the 19th Century and the loss of American chestnut to chestnut blight in the first part of the 20th Century. Therefore, the response of future forests to changes in temperature and rainfall might now depend on these novel tree-shrub interactions .
R. maximum hampers succession by forming a dense subcanopy layer that greatly limits light reaching the forest floor and reduces soil moisture and temperature. These changes impede seed germination and seedling survival. In addition, rhododendron leaves that fall to the ground create a thick organic soil layer that decomposes very slowly. This affects soil chemistry, specifically availability of the key nutrient nitrogen.
The rhododendron shrubs in the region are younger than the deciduous trees now making up the canopy above them (Dharmadi, Elliott and Miniat 2019). The dense rhododendron stands resulted from the widespread mortality of American chestnut (Castanea dentata) in the early 20th century and of hemlock in the first years of the 21st Century. What’s more, even the mature deciduous trees appear to be suppressed by dense rhododendron stands. Canopy trees above rhododendrons are on average 6m shorter than those growing on sites without rhododendron thickets (Dharmadi, Elliott and Miniat 2019). In fact, by 2014, 10% of standing trees other than hemlocks had died. The tree suffering the highest level of mortality was flowering dogwood (Cornus florida). The authors do not mention a probable factor, the introduced disease dogwood anthracnose.Other species experiencing high levels of mortality are not, to my knowledge, under attack by non-native pests, so their demise seems more clearly linked to resource competition with rhododendron. These were striped maple (Acer pennsylvanicum), pitch pine (Pinus rigida), witch hazel (Hamamelis virginiana), and that staple of New England aftermath forests, black birch (Betula lenta).
Dharmadi, Elliott and Miniat (2019) suggested that managers should step in to increase recruitment in both understory and overstory layers. They proposed active management: removing rhododendrons and the soil organic layer. USFS scientists are applying these ideas experimentally at the Coweeta research station. I am unclear as to whether there is one study or more. In any case, rhododendronplants have been removed with the goal of restoring vegetation structure and composition – presumably both understory plant diversity and recruitment of tree species capable of growing into the canopy. In at least some cases, the rhododendron removal is followed by prescribed fire. One study is looking also at whether this action increased water yield.
Apparently this lack of tree regeneration is extensive – although published data are not easily accessible. Staff of the North Carolina Hemlock Restoration Initiative report they encounter similar issues (O.W. Hall, Hemlock Restoration Initiative, pers. comm.)
Several experiments have demonstrated that even in the southern Appalachians, where there are abundant moisture and rainfall, the trees and shrubs compete for water and other nutrients. However, Dharmadi et al. (2022) found that removal of the rhododendron shrub layer is unlikely to significantly alter streamflow, atr least during the growing season. In winter, when deciduous trees lack leaves, reduction in interception of precipitation might result in increased streamflow (Dharmadi et al. 2022). I ask whether increasing stream flow in winter is a goal? I thought the concern was stream flow levels in summer.
Nor is removal of the rhododendron shrub layer likely to alter stream chemistry during the growing season.
Removal of living Rhododendron and leaf litter apparently can help restore forest structure through improving tree seedling survival and recruitment as well as increasing growth of established trees.
Removing Privet
However, other management actions might bring about desired changes more effectively or broadly. Specifically Dharmadi and colleagues mentioned removal of privet (Ligustrum) – a very widespread invasive shrub in forests of the Southeast. (Fifteen years ago it was estimated that just one privet species, Chinese privet, occupied more than a million hectares in 12 southeastern states [Hanula 2009].)
Chinese privet
I ask also whether prescribed fire to remove the rhododendron-dominated soil organic layer is useful. Dharmadi and colleagues found that such fires reduced leaf litter temporarily, but annual leaf-fall replaced the litter layer the next year, so this management effort is unlikely to affect plot evapotranspiration rates.
Supporting Pollinators
Another study (Ulyshenet al. 2022) examined whether removing rosebay rhododendron would benefit bees and other pollinators. They found that removal of Rhododendron alone (without fire) did not dramatically improve pollinator habitat in the southern Appalachians. In fact, about a quarter of the bee species studied visited R. maximum flowers and might decline if the shrub’s population is reduced. Ulyshen and colleagues suggest that some factors that correlate with fire severity probably promotes growth of insect-pollinated plants. They suggest specifically the greater presence of downed woody debris, which provides nesting sites and other resources used by insects. They recommended creation of open areas to support wildflowers as a more effective way to benefit bees in this region. Again, rhododendron removal pales in effectiveness compared to eradication of privet.
SOURCES
Dharmadi, S.N., K.J. Elliott, C.F. Miniat. 2019. Lack of forest tree seedling recruitment and enhanced tree and shrub growth characterizes post-Tsuga canadensis mortality forests in the southern Appalachians. Forest Ecology and Management 440 (2019) 122–130.
Dharmadi, S.N., K.J. Elliott, C.F. Miniat. 2022. Larger hardwood trees benefit from removing Rhododendron maximum following Tsuga canadensis mortality. Forest Ecology and Management
Hanula, J.L., S. Horn, and J.W. Taylor. 2009. Chinese Privet (Ligustrum sinense) Removal and its Effect on Native Plant Communities of Riparian Forests. Invasive Plant Science and Management 2009 2:292–300.
Plotkin, A.B., A.M. Ellison, D.A. Orwig, M.G. MacLean. 2024. Logging response alters trajectories of reorganization after loss of a foundation tree species. Ecological Applications. 2024;e2957.
Ulyshen, M., K. Elliott, J. Scott, S. Horn, P. Clinton, N. Liu, C.F. Miniat, P. Caldwell, C. Oishi, J. Knoepp, P. Bolstad. 2022. Effects of Rhododendron removal and prescribed fire on bees and plants in the southern Appalachians. Ecology and Evolution. 2022;12:e8677.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
Reminder: beech leaf disease (BLD) came to attention in 2012 near Cleveland. It has since spread rapidly to the East and more slowly to the North, South, and West. It has been detected in 15 states and the Province of Ontario. The disease is caused by the foliar nematode Litylenchus crenatae mccannii (Lcm). Damage to the leaves can significantly reduce the tree’s ability to photosynthesize, resulting in a progressive depletion of carbohydrate reserves following successive years of infection. Scientists continue efforts to determine how it is spread and the extent of tree mortality.
Last summer I blogged about an Integrated Pest Management (IPM) strategy developed by Bartlett Tree Research Laboratories – the research arm of Bartlett Tree Experts – to treat individual beech trees afflicted with beech leaf disease (BLD). This treatment relied on a foliar spray. The challenge is that the entire canopy must be sprayed; this is difficult for large trees. Also, some trees cannot be sprayed because of their proximity to water bodies or other issues. For these large trees, Bartlett sought to develop a systemic treatment that can be applied as a drench or root flare injection.
photo by Matt Borden, Bartlett, via Flickr
I rejoice to tell you that Bartlett has now confirmed effectiveness of a systemic root flare injection treatment. Again, the project is led by Dr. Andrew Loyd and Dr. Matthew Borden. The full citation for their publication is at the end of this blog.ph
This second treatment utilizes Thiabendazole (TBZ), which has a long history of use in arboriculture to manage Dutch elm disease and sycamore anthracnose. In addition to being a fungicide TBZ is also a potent nematicide.
Bartlett tested the efficacy of TBZ by monitoring 62 symptomatic American beech (Fagus grandifolia) trees across three sites comprising natural mixed hardwood forests with beech as a dominant species. Half of the trees were injected with TBZ, and the rest were monitored as non-treated controls. In the late winter after applying the injection, the researchers sampled twigs from all the trees and counted the number of nematodes in late-season dormant buds, where most of the damage occurs. They also quantified canopy density and BLD symptom expression before treatment and around 11 months after the tree injections the following year, to gauge year-over-year change. At one site, in Ohio, the trees were assessed again in the second season, or 22 months, after the initial injection. They also assessed damage to the root flare caused by the injection process.
Bartlett’s researchers found that at both 11 and 22 months after treatment, injected trees had significantly better visual ratings of canopy condition and lower numbers of Lcm in dormant buds. The untreated controls continued to have high disease severity and large numbers of nematodes in their buds.
Detailed Results
At the time of the initial inventory and treatment, about 65% of the canopies of beech trees at the two Ohio sites displayed foliar BLD symptoms. The proportion was lower at the New Jersey site, where BLD has been present only a season or two before treating – 42%.
During the first growing season post-treatment, the percent of the symptomatic beech canopies at the two Ohio sites fell by 70 – 85%. At the site where trees were evaluated again the second season (22 months) post-treatment, the percent of the canopy exhibiting leaf symptoms continued to decline. The scientists hypothesize that this continuing decrease could be due to TBZ residues being translocated to new leaves and buds, or to a reduction in local inoculum sources within the individual trees and surrounding forest due to treating a significant portion of the community.
At the New Jersey site, where injection was performed later in the season, the percent of the canopy exhibiting leaf symptoms increased by 66%. However, by another measure – percent of canopy with fine twig dieback – these trees improved by 71% while on untreated trees twig dieback increased by 95% and were already experiencing severe canopy loss.
New Jersey site contrasting treated & untreated trees; photo by Matt Borden, Bartlett
On average, there were significantly fewer Lcm in dormant bud tissues in treated trees compared to the untreated control trees. At the two sites in Ohio, the reductions were by 86% and 99%. At the New Jersey site, the decline was not as great, but still encouraging: 70%.
These results suggest that one treatment can substantially reduce symptoms. Scientists now need to determine at what point BLD symptoms return to damaging levels at both “low” & “high” concentrations of thiabendazole in order to determine retreatment intervals and expectations.
While the disease severity (measured by the percent canopy displaying BLD leaf symptoms) of all trees increased at Hillsborough, the canopies of trees injected with the “low rate” of TBZ was significantly better than those of the untreated trees. This was because of a significant reduction in fine twig dieback in the former as opposed to a significant increase in fine twig dieback in untreated controls. Fine twig dieback symptom expression is presumed to be associated with bud abortion caused by Lcm.
The New Jersey treatments occurred at the end of August. The scientists think that this period might follow rather than precede dispersal of many nematodes from the leaves to the buds, as evidenced by the reduced but still substantial numbers of nematodes found in the buds.
While there was some damage visible at injection sites, the Bartlett team considers the frequency of these symptoms to be low. Cracking of the bark was seen on 19% of injected trees; evidence of fluxing was present on 12%. Injection sites were closing rapidly at the site reviewed after 22 months post-treatment. Additionally, based on observations made during this study, they believe that cracking can be further reduced.
Loyd et al. conclude that TBZ injection is an effective treatment option for large beech (> 25-cm dbh) where full coverage sprays with fluopyram are difficult, or for trees growing near water, or where pesticide drift may be of concern.
a forest in Northern Virginia dominated by beech; photo by F.T. Campbell
While this treatment can be used in natural landscapes, treatments of whole forests will probably not be feasible due to the cost. Scientists continue investigating whether some combination of silvicultural practices such as reduction in stand density and with pesticide application of select mature beech might prove effective. In fact, scientists are establishing new plots this year to test a silviculture management approach in forests of Pennsylvania and Rhode Island where BLD is prevalent.
SOURCE
Loyd, A.L., M.A. Borden, C.A. Littlejohn, C.M. Rigsby, B. Brantley, M. Ware, C. McCurry, & K. Fite. 2025. Thiabendazole as a Therapeutic Root Flare Injection for Beech Leaf Disease Management Arboriculture & Urban Forestry 2025 https://doi.org/10.48044/jauf.2025.007
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
The pest alert system “PestLens” has again alerted us to plant pests in Europe or Asia that feed on species closely related to tree species native to North American forests. Two of the insects named in the alert apparently pose a hazard to icons of the forests of America’s Pacific coast forests, giant sequoia and redwood.
I hope APHIS is using this information to alert port and on-the-ground staff and perhaps initiating more in-depth risk assessments.
The posting on February 27, 2025 reported that cotton jassid, Jacobiasca lybica (Hemiptera: Cicadellidae), affects not just cotton and citrus but also Cupressus sempervirens (Mediterranean cypress) [Cupressaceae]. More than a dozen North American trees species are in this family, including
Sequoiadendron giganteum or giant sequoia. Giant sequoia is listed as an endangered species by the IUCN with fewer than 80,000 remaining in its native California.
Chamaecyparis thyoides and C. lawsoniana (Port-Orford cedar). Port-Orford cedar has been decimated in its native range by an introduced pathogen, Phytopthora lateralis. A major breeding effort has developed trees that are resistant to the pathogen; they are now available for people to plant.
Thuja occidentalis, also known as northern white-cedar, eastern white-cedar, or arborvitae,
Taxodium ascendens, also known as pond cypress
several Juniperus
Hesperocyparis macrocarpa also known as Cupressus macrocarpa, or the Monterey cypress. NatureServe ranks the cypress as GI – critically imperiled.
Cotton jassid been reported from several countries in Europe, Africa, and the Middle East.
China has reported the existence of a previously unknown bark beetle species, Phloeosinus metasequoiae (Coleoptera: Curculionidae). It was found infesting Metasequoia glyptostroboides (dawn redwood) trees in China. Affected trees exhibited reddened leaves and holes and tunnels in branches.
China has also discovered a several new hosts utilized by the fungus Pestalotiopsis lushanensis (Sordariomycetes: Amphisphaeriales). Formerly known to infect tea (Camellia sinensis) and several other plant species, P. lushanensis has now been found shoot causing blight and leaf drop on a conifer, deodar cedar (Cedrus deodara) and leaf spots on an angiospermwith congeners in North America — the rare Chinese species, Magnolia decidua. There are eight species of Magnolia native to North America.
Magnolia grandiflora; photo by DavetheMage via Wikimedia
APHIS’ ability to respond to alerts remains uncertain.
The agency’s probationary employees have been fired – just as at other agencies. APHIS staff were prohibited from participating in last week’s annual USDA Invasive Species Research Forum – the 33rd such meeting. The bird flu emergency is demanding all the attention and funds.
So – how can the rest of us fill in?
At the USDA Research Forum I again presented a poster urging greater attention to tree-killing pathogens. Scientists have made considerable progress in identifying factors that indicate whether a non-native insect might pose a significant threat (see blogs on conifer and deciduous species; more to come!). However, USDA had not funded a similar effort to improve understanding of pathogens. The most promising strategy so far are sentinel plantings. However, these systems have weaknesses; I will blog in the near future about another analysis.
I propose that APHIS start by working with independent scientists to determine the actual, current level of pathogens associated with various types of incoming goods. Contact me directly if you wish to read the text of my poster.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/










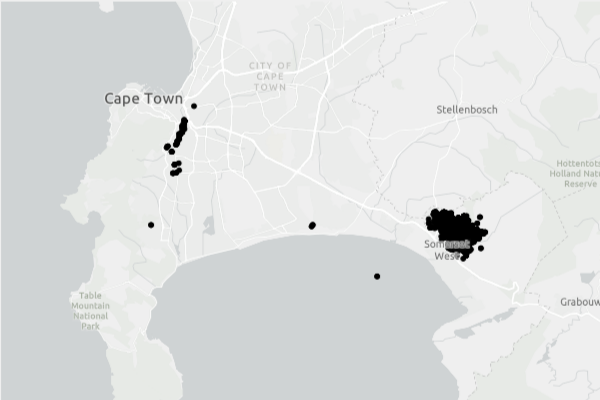
























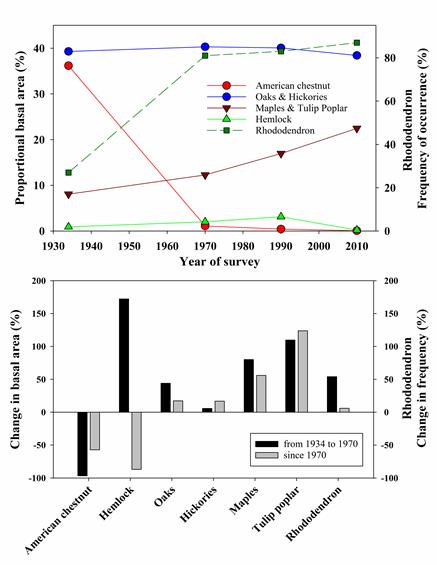{kind=link}