Scientists in New Zealand are saying explicitly that a forest’s unique mixture of species matters when considering the future. This mixture is the result of the forest’s evolutionary history. Losing members of the biological community reduces the forest’s ability to respond to current and future stresses – its resilience.
New Zealand’s forests are part of the broader legacy of the ancient supercontinent of Gondwanaland – the island nation’s plants have close relatives in South America, the Pacific Ocean islands, and Australia. Still, these forests are unique: 80% of New Zealand’s plant species are endemic. The forests are also species-rich. The warm temperate evergreen rain forests of the North Island are home to at least 66 woody plant species that can reach that reach heights above six meters (Simpkins et al. 2024).
These forests have been severely changed by human activity. In just ~ 750 years people have cut down approximately 80% of the original forest cover! (Simpkins et al. 2024) Of the eight million hectares of surviving native forest, a little over five million hectares is managed for the conservation of biodiversity, heritage, and recreation. Another 2 million hectares are plantations of non-native species.

All these forests are challenged by introduced mammals – from European deer to Australian possums. Climate change is expected to cause further disturbance, both directly (through e.g., drought, extreme weather) and indirectly (e.g., by facilitating weed invasion and shifting fire regimes) (Simpkins et al. 2024).
Pathogen threats are also common threats to the native trees of the Pacific’s biologically unique island systems. For example, Ceratocystis lukuohia and C. huliohia (rapid ‘ōhi‘a death, or ROD). The latter is killing ‘ōhi‘a (Metrosideros polymorpha) on the Hawaiian Islands. More than 40% of native plant species in Western Australia are susceptible to Phytophthora cinnamomi. Here I focus on two pathogens, kauri dieback and myrtle rust, now ravaging New Zealand’s native flora. No landscape-level treatment is available for either pathogen.
When considering this suite of challenges, Simpkins et al. focus on these two pathogens’ probable impact on forest carbon sequestration. They worry in particular about erosion of the forests’ resilience due to loss of “ecological memory” – the life-history traits of the species (e.g., soil seed banks) and the structures left behind after individual disturbances.

Kauri Dieback
The causal agent of Kauri dieback, Phytophthora agathidicida, is a soil-borne pathogen that spreads slowly in the absence of animal or human vectors. The disease affects a single species, Agathis australis (kauri, Araucariaceae). However, kauri is a long-lived, large tree that is a significant carbon sink. It probably modifies local soil conditions, nutrient and water cycles, and associated vegetation. Also, kauri has immense cultural significance.
Simpkins et al. note that kauri dieback threatens stand-level loss of A. australis – that is, local extinctions. In the absence of disturbance Kauri trees can grow to awe-inspiring size. In the 19th Century, before widespread logging, some were measured at 20 meters or more in circumference. Consequently, kauri dieback might cause a decline in aboveground live carbon storage of up to 55%. This loss would occur over a period of hundreds of years, not immediately.
Huge kauri are not likely to be replaced by other long-lived emergent conifers (based on an analysis of one species, Dacrydium cupressinum). Instead, kauri are probably going to be replaced by late-successional angiosperms. The authors discuss the ecological implications for levels of carbon storage and proportions of trees composed of Myrtaceae – exacerbating damage caused by myrtle rust (see below).
The expectation of Simpkins et al. that kauri will suffer at least local extinctions is based on an assumption that no kauri trees are resistant to the pathogen. Fortunately, this might not be true: different Agathis populations show various levels of tolerance to Agathis dieback. Identification and promotion of some levels of resistance could enable A. australis to retain a diminished presence in the landscape.
However, Lantham, et al. make clear that containing kauri dieback remains “challenging,” despite its discovery nearly 20 years ago (in 2006). Scientists and land managers have little information on the distribution of symptomatic trees, much less of the pathogen itself. This means they don’t know where infection foci are or how fast the disease is spreading.
As is often true, the pathogen is probably present in a stand for years, possibly a decade or more, before symptoms are noticed. This means that the current reliance on public reports of diseased trees, or targetting surveillance on easy-to-access sites (e.g., park entrances and along existing track networks), or at highly impacted areas readily identified through aerial methods, fails to detect early stages of infection. Indeed, it seems probable that P. agathidicida had been present in New Zealand’s ecosystems for decades before its formal identification.
The Waipoua forest is one of the largest areas of forest with old kauri stands in the country. A new analysis of aerial surveys done between 1950 and 2019, shows how the forest is changing. The number of dead trees increased more than four-fold and the number of unhealthy-looking trees increased 16-fold over these 70 years. Kauri dieback is now widespread in this forest, especially in areas near human activities like clearing for pasture or planting commercial pine plantations).
Lantham et al. have developed a model which they believe will help identify areas of higher risk so as to prioritize surveillance and inform responses. These could delimit the disease front and help implement quarantines or other measures aimed at limiting the spread of P. agathidicida to uninfected neighboring sites.
I hope New Zealand devotes sufficient resources to expand surveillance and management to levels commensurate with the threat to this ecologically and culturally important tree species.

Myrtle Rust
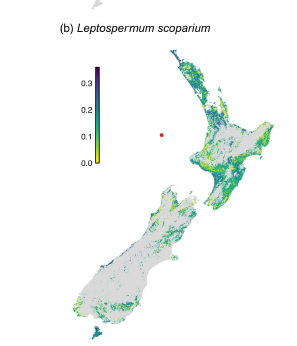
Myrtle rust is a wind-borne disease that affecting numerous species in the Myrtaceae, including some of the dominant early successional species (e.g., Leptospermum spp.). Simpkins et al. expect that myrtle rust might hasten the decline of two such tree species (L. scoparium and Kunzea ericoides). However, these trees’ small size and rapid replacement by other species during succession minimizes the effect of their demise on carbon storage.
Because I am concerned about the irreplaceable loss to biodiversity, I note that Simpkins et al. also feared immediate threats to some trees in the host Myrtaceae family, specifically highly susceptible species such as Leptospermum bullata.
As I reported in a recent blog, a second group of scientists (McCarthy et al.) explored the threat from myrtle rust more broadly. Austropuccinia psidii has spread through Myrtaceae-dominated forests of the Pacific islands for about 20 years.
Trees in the vulnerable plant family, Myrtaceae, are second in importance (based on density and cover) in New Zealand’s forests. Successional shrub communities dominated by the two species named above, Kunzea ericoides and Leptospermum scoparium, are widespread in the northern and central regions of the North Island and in northeastern and interior parts of the South Island. These regions’ vulnerability is exacerbated by the area’s climate, which is highly suitable for A. psidii infection (Simpkins et al. 2024).
McCarthy et al. concluded that if Leptospermum scoparium and Kunzea ericoides prove to be vulnerable to myrtle rust, their loss would cause considerable change in stand-level functional composition across these large areas. They probably would be replaced by non-native shrubs, which are already common on the islands. Any resulting forest will differ from that formed via Leptospermeae succession.
These authors also worry that the risk to native ecosystems would increase if more virulent strains of the myrtle rust pathogen were introduced or evolved. They note that A. psidii is known to have many strains and that these strains attack different host species.
SOURCES
Latham, M.C., A. Lustig, N.M. Williams, A. McDonald, T. Patuawa, J. Chetham, S. Johnson, A. Carrington, W. Wood, and D.P. Anderson. 2025. Design of risk-based surveillance to demonstrate absence of Phytophthora agathidicida in New Zealand kauri forests. Biol. Invasions (2025) 27, no. 26
McCarthy, J.K., S.J. Richardson, I. Jo, S.K. Wiser, T.A. Easdale, J.D. Shepherd, P.J. Bellingham. 2024. A Functional Assessment of Community Vulnerability to the Loss of Myrtaceae from Myrtle Rust. Diversity and Distributions, https://doi.org/10.1111/ddi.13928
Simpkins, C.E., P.J. Bellingham, K. Reihana, J.M.R. Brock, G.L.W. Perry. 2024. Evaluating the effects of two newly emerging plant pathogens on North Aotearoa-New Zealand forests using an individual-based model. Ecological Modelling, www.elsevier.com/locate/ecolmodel
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
or