Reminder: Friday is the deadline for commenting on APHIS’ draft environmental assessment for the Asian longhorned beetle eradication program in South Carolina. Comments should be submitted at https://beta.regulations.gov/commenton/APHIS-2020-0086-0001
The Center for Invasive Species Prevention submitted comments that supported the eradication effort because of the well-documented threat that the ALB poses to the forests of North America. We also supported the preferred alternative in the EA.
However, we found the environmental assessment (EA) to be deficient in several ways:
the EA does not identify the host species present in the program area – not even of the 5,800 trees inspected by the program as of mid-August.
the EA provides no estimate of the proportion of deciduous trees and shrubs in the area that are host species. Conifers dominate the area. This means that any fauna dependent on deciduous trees and shrubs for food and shelter already contend with limited resources. Consequently, while we concur with the EA that any impacts will be localized, they might be exacerbated by the relative rarity of hardwood species in the local area. It is particularly important that the EA address this question since the Programmatic EIS was written under the assumption that forests at risk to the ALB are like those in the Northeast and Midwest, where hardwoods dominate.
Without knowing the proportion of deciduous flora comprised of host species, no one can evaluate the amount of wildlife food that could be removed or treated by pesticides. Some wildlife species are potentially vulnerable, including those that feed on pollen and nectar (i.e., bees and other pollinators) and those that feed on insects and other invertebrates. The latter include two species listed federally as threatened species: the frosted flatwood salamander (Ambystoma cingulatum) and northern long-eared bat (Myotis septentrionalis). Also vulnerable are birds, 96% of which feed their young on insects and other invertebrates. I worry about sublethal effects and possible bioaccumulation. Aquatic organisms, especially invertebrates, might also be affected.
The information gaps in the EA highlight weaknesses in the Programmatic EIS, on which it relies. The most important gap is the dearth of pesticide dose/mortality data for terrestrial amphibians. Apparently, EPA has not required such studies before approving pesticides.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
APHIS has apparently passed up an opportunity to pressure China to clean up its wood packaging – although China ranks among the countries that most often violates ISPM#15 and sends wood packaging infested by quarantine pests. (See the blogs under the category “wood packaging” on this site.)
In May, a large delegation of APHIS employees met (virtually) with an equally large delegation of its Chinese counterpart to negotiate “technical protocols” linked to the Phase 1 trade agreement with China. The focus of the negotiations was on Chinese phytosanitary barriers that block exports of US products to China.
The two countries have now signed technical protocols to allow the United States to export to China a wide range of commodities estimated to be worth between $700 million and $760 million annually when the agreement is fully implemented. These commodities include barley for processing, hay, some fruits (blueberries, avocados, nectarines), almond meal, and chipping potatoes.
Some of the agreements cap years of effort. The example cited is chipping potatoes. Negotiations continue on some other U.S. exports to China, including logs.
An article in APHIS’ online newsletter reports that “On the import side, we are working on the requirements for China’s requested commodities….” Presumably these would be exports to the U.S. The examples listed were all fruits.
US & Chinese delegations (APHIS photo)
I inquired whether wood packaging was part of the negotiation.
Andrea B. Simao, Assistant Deputy Administrator and Director of PPQ’s Phytosanitary Issues Management unit, replied that SWPM was not raised “since there has [sic] not been significant issues.” Instead, she detailed efforts in the ongoing negotiations to persuade China that U.S. phytosanitary treatments are sufficient to control various pathogens in logs: oak wilt, phosphine on conifers, pinewood nematode.
Apparently the focus was fully on US exports and nobody raised US concerns about the risks of imports from China. This approach fits the Administration’s emphasis on exporting agricultural commodities to China. However, this is not reality. Over the past five years, I have frequently cited USDA’s own data – which demonstrate the likelihood that wood packaging will transport tree-killing pests from China to the U.S.
APHIS PPQ Deputy Administrator Osama El-Lissy & Chinese counterpart Li Jainwei sign agreement (APHIS photo)
Please inform your Member of Congress and Senators (or candidates for House or Senate) about how you feel about this failure of USDA to protect America’s natural resources. We must raise the political heat in order to pressure USDA into placing as high a priority on protecting US natural resources as it does on supporting agricultural exports.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
U.S. phytosanitary policy is set by politicians – the Secretary of Agriculture, trade officials, and members of the House and Senate. Elected or appointed state officials determine how aggressively trees are protected in their jurisdictions. To fix the problems, those politicians need to hear from those of us who know about the pest risk associated with wood packaging and other imports.
Politics is how our country makes important decisions. And in politics, the squeaky wheel gets the grease.
Election seasons provide opportunities to raise issues. Politicians pay more attention to constituents’ concerns when they are courting our votes.
Further, if new people take up positions in January (whether elected or appointed), they will be more open to learning about issues new to them than were the people who have occupied an office for some time.
These messages need to be repeated periodically. Proctor and Gamble does not make its profits by asking us to buy their toothpaste once a year. We cannot duplicate a major corporation’s advertising budget – but we can speak up!
Tell your member of Congress and senators that you are worried that our trees are still being put at risk by insects arriving in wood packaging or diseases being spread by shipments of plants. Ask them to urge theUSDA Secretary to take action to curtail introductions of additional tree-killing pests.
Ask your friends and neighbors to join you in communicating these concerns to their Congressional representatives and senators.
If you are a member of an association – a scientific or professional society, an environmental advocacy group, a homeowners’ association – ask your association and fellow members to join you in communicating these concerns to their Congressional representatives and senators.
Write letters to the editors of your local newspaper or TV news station.
What should we say?
Our goal should be to hold foreign suppliers responsible for complying with ISPM#15. Here are five pieces of a comprehensive approach. It is best to advocate for all. However, if you feel more comfortable focusing on one or two specific actions, please do so!
1) One approach is to penalize violators. APHIS should:
Fine an importer for each new shipment found to be out of compliance with ISPM#15 in those cases where the foreign supplier of that shipment has a record of repeated violations.
Prohibit imports in packaging made from solid wood (boards, 4 x 4s, etc.) from foreign suppliers that have a record of repeated violations.
Allow continued imports from those same suppliers as long as they are contained in other types of packaging materials, including plastic, metals, fiberboards.
APHIS has the authority to take these action under the “emergency action” provision (Sec. 5.7) of the World Trade Organization Agreement on Sanitary and Phytosanitary Standards. (See a lengthy discussion of the SPS agreement in Chapter III of Fading Forests II, available here.) http://treeimprovement.utk.edu/FadingForests.htm
USDA and CBP should take other steps to help importers comply with ISPM#15.
USDA should also step up efforts to help U.S. importers to determine – and then use – those foreign suppliers of wood packaging and dunnage have good compliance records.
APHIS should join the DHS CBP in providing incentives to importers to join an expanded Customs-Trade Partnership Against Terrorism program (C-TPAT) that would require participants to assume full responsibility for ensuring that their packaging complies with IPPC standards.
The Government should strengthen underlying regulations.
Once a new president is elected, urge him to instruct the Office of Management and Budget to allow APHIS to finalize regulations – proposed more than five years ago! – that would apply ISPM#15 to wood packaging used in trade between the US and Canada. (Canada has been ready to adopt this measure for several years.)
USDA needs to understand the “approach rate” of pests in wood packaging in order to identify and fix weaknesses in its policies. To reach this understanding, APHIS should authorize Robert Haack to repeat the study documented in Haack et al. (2014). Furthermore, APHIS should collaborate with foreign counterparts to determine the relative importance of possible causes of the persistent pest presence problem – fraud, accidental misapplication of treatments, or other failures of treatment. Once the study has been completed, APHIS and its colleagues should work through the IPPC to fix the problems.
There are also recommendations of the Tree-Smart Trade program at www.tree-smart-trade.org Tree-Smart also has a Twitter account: @treeSMARTtrade
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Ash forest in Michigan killed by emerald ash borer photo by Nate Siegert
As I reported in December, the USDA Forest Service Forest Health Management program has been issuing reports on the status of pest invasions, based on annual national Insect and Disease Surveys (IDS). This survey is carried out by low-level aerial surveys of statistically valid geographic units. (For a discussion of quadrat establishment, see the Introduction to the 2019 report here.
Here I compare the pest situation as it appeared in two recent years, based on these surveys: 2017 and 2018.
Of course, two years is too short a time to see trends. The second report provides USFS scientists attempt to provide context by analyzing 20 years of data (Chapter 6 of the 2019 report).
These reports are important sources of data on the status of non-native pests in our forests, but I raise several caveats that necessitate our continuing to rely on additional sources, some of which I have cited in previous blogs. I will address these in greater detail below. But in brief, these caveats are:
1) Major pests – both in extent and damage caused – are not adequately included (see below).
2) The survey technique resulted in delays in detecting mortality. It cannot be used as an early detection tool.
3) There is little analysis of the statistics cited.
Why Stakeholders Cannot Rely on These Two Survey Reports Alone
I hope stakeholders, analysts, and decision-makers will utilize the full range of reports and other data to evaluate the presence and impacts of various invasive pests and pathogens. Even the reports’ authors advise decision-makers to use other forest health indicators in addition to this report. It is not the “fault” of the authors that these reports cannot serve all needs. However, we all need to be aware this caveat.
1. Important Pests Not Adequately Included.
Many of the most damaging non-native pests caused extensive damage before these surveys were initiated. These include white pine blister rust, chestnut blight, Dutch elm disease, butternut canker, Port-Orford cedar root disease, European gypsy moth, and balsam woolly adelgid on Fraser fir. [For further discussion, see pest write-ups here and my earlier blogs reviewing 30 years of engagement and survey data from Shenandoah National Park.]
2. Major Damaging Pests Are Poorly Captured by the Survey Method.Morin found that non-native pests had caused a five percent increase in mortality nation-wide – as measured by tree volume. Three of the species suffering the highest rates of this “exacerbated mortality” are redbay, ash, and hemlock. The current studies’ authors concede all three are poorly detected by the survey methods. The aerial survey techniques are ill-suited to detect mortality of tree species that grow scattered throughout a diverse forest. Other seriously damaging pests that are poorly detected by aerial surveys are Dutch elm disease, white pine blister rust, and thousand cankers disease. Again, see the species write-ups here to be reminded about how great the pest’s impact are. Also review my earlier blog reviewing 30 years of my engagement with the issue and survey data from Shenandoah National Park.
dead redbay in woods photo by F.T. Campbell
Given these caveats above, what do these studies show us?
Areas surveyed.
Since 1999, the annual Insect and Disease Survey (IDS) has covered on average 266,655,000 ha, although the area varied from a maximum of 320,712,000 ha (in 2007) to a minimum of 202,170,000 ha (in 2017), then increased to 211,34,000 ha in 2018. For reasons not clear to me, the proportion of forested area in the lower 48 states surveyed actually declined from 2017 to 2018 – from 55.1% to 46.6%. In Alaska, the 2017 surveys covered about 7.3% of the total forested area but 12.7% of the forested area in 2018. In Hawai`i, the survey covered about 80.1% of the forested area in 2017, but only 69.4% of the State’s total tree canopy area in 2018.
Tree-Killing Insects and Pathogens Found.
The 2017 survey identified 63 mortality-causing agents and complexes that cumulatively affected 3.27 million ha in the lower 48 states – about 1.3% of the total 252 million ha of forested land in these states. Of these agents of tree mortality, 23 were detected killing trees on areas totaling larger than 5,000 ha each.
The 2018 survey identified seven fewer mortality-causing agents – only 56. These agents were detected on about 2.13 million ha across the lower 48 (slightly less than the combined land area of New Jersey and Rhode Island), or about 0.8% of the total forested area. Of the total, 22 agents were detected killing trees on areas totaling larger than 5,000 ha each.
The more recent report does not discuss these declines from the 2017 findings or whether they might be related to the smaller percentages of forested areas covered by the aerial survey in 2018.
In both 2017 and 2018, as well as in the 20-year trend analysis (Chapter 6 of the report published in 2020), overall mortality is greatest in the West due to the impact of several native western bark beetles. Overall mortality rates in other regions was considered low, despite severe impacts of some non-native species.
The 2017 survey found that the emerald ash borer (EAB) was the most widespread single agent, causing measurable tree mortality on 1.42 million ha. In 2018, the area of EAB damage was reduced by 76% — to just 338,000 ha. (Still, the latter figure represented 15.8% of the total area displaying mortality). These figures were probably underestimates – especially in areas outside North Central Region – because EAB is one of the pests poorly detected by the aerial survey technique – at least when ash are growing scattered in a diverse forest ecosystem. Although the USFS report doesn’t say so, this decline probably reflected the collapse of dead ash trees and reduction in numbers of still-alive but vulnerable ash trees as the EAB invasion wave matured.
The reports document a huge increase in mortality attributed to the sudden oak death pathogen between 2017 and 2018. (SOD is the only widespread non-native agent of mortality on the West coast.) The area affected increased nearly seven-fold – from 6,335 ha in the 2017 survey to 42,771 ha in the 2018 survey. This equated to causing mortality on 3.9% of the total Regional mortality area. This finding parallels finding reported by the California Oak Mortality Task Force. Note that SOD-related mortality was not detected by the aerial surveys until 2008 – 13 year or more after scientists working on the ground detected the presence of the then-unknown pathogen in the forests of California.
tanoak killed by Phytophthora ramorum on Big Sur peninsula photo by Matteo Garbelotto
Another alarming increase detected in 2018 was that of balsam woolly adelgid (BWA) in the Interior West. BWA-caused mortality was evident on 44,000 ha – 8.4% of the total area with mortality. BWA impacts were especially severe in central Idaho. There are several native pests in the same area. (See BWA write-up here.) BWA was first detected in Idaho in 1983 – 35 years earlier.
Geographic Hot Spots
The EAB caused the principal hot spots in the East. In the USFS North Central Region, 91% of the area suffering tree mortality in 2017 was attributed to the EAB. The EAB was also causing mortality across 10,346 ha (16% of the total areas suffering mortality) in the Northeast (especially Connecticut) and more than 5,000 ha in the South (especially Kentucky). (See my December blog for a discussion of the puzzling situation in several Great Plains ecoregions, where mortality was attributed largely to drought rather than either EAB or Dutch elm disease.)
Another mortality agent in the Northeastern Region was the European gypsy moth. In 2018, it was responsible for tree death on 31% of the total 70,000 ha affected area.
Another hot spot was in Hawai`i. In 2017, about 37,000 ha of mortality was detected. By 2018, the dead zone had increased to 46,000 ha – despite a 12% decrease in the area surveyed. In both years, the data collectors officially called the cause unknown. The reports’ authors stated that the probable cause was they rapid ‘ōhi‘a death fungi. (I ask why the surveyors did not state the cause since rapid ‘ōhi‘a death was identified on the Big Island in 2014.)
rapid ‘ōhi‘a death photo by J.B. Friday
Other non-native pests that affected more than 5,000 ha in the lower 48 states in 2017 were the BWA outbreak in the Northeast (20,758 ha, primarily in Maine); beech bark disease (12,222 ha, primarily in the North Central Region), and oak wilt (9,573 ha, primarily in the North Central Region and Texas).
In the Southern Region, mortality agents were detected on 1% or less of the forested area in 2017. In 2018, EAB-caused mortality was detected on 9.7% of the total 13,000 ha area experiencing mortality. An earlier gypsy moth outbreak had apparently calmed. I remind you that the report authors have conceded that laurel wilt and hemlock woolly adelgid are poorly detected by the survey technique.
eastern hemlock in Shenandoah National Park (Virgina)
DEFOLIATORS
The 2017 survey detected defoliation caused by 50 agents and complexes across the lower 48. These impacted 2.3 million ha. The 2018 survey identified six more defoliation agents and complexes (56), but they affected a much smaller area – about 1.72 million ha.
Unsurprisingly, the most widespread was the European gypsy moth. The data demonstrated the gypsy moth’s boom/bust cycle. In 2017, gypsy moth impacts were detected on 39% of the total forested area of the lower 48 states (913,000 ha) in 2017. By 2018, the total area affected by the gyspy moth had fallen to 156,000 ha.
In both years, gypsy moth defoliation was particularly severe in the Northeast Region. In 2018, a second non-native species, browntail moth (Euproctis chrysorrhoea) was also causing severe defoliation in the region, primarily in Maine.
In the South, the European gypsy moth affected only 5.8% of the total area of defoliation; native defoliators predominated.
The report does not discuss the relationship between gypsy-moth caused defoliation and mortality. True, there must be repeated defoliations to cause tree mortality.
Other non-native defoliation agents affecting more than 5,000 ha in the lower 48 in 2017 were the larch casebearer (25,891 ha in the North Central Region and another 7,400 ha in the West Coast Region); and winter moth (12,760 ha in the Northeast Region). Is the decline of winter moth in 2018 data related to introduction of a biocontrol agent?
In 2018, the Larch casebearer (Coleophora laricella) continued to be a significant defoliator, affected 3.1% of the North Central Region’s total defoliated area. A new agent, the balsam woolly adelgid (BWA), was detected defoliating firs in the West Coast Region. The affected area was 15,000 ha – 5% of the total affected area Again, there was no discussion of as to whether defoliation precedes mortality. Admittedly, the progression of BWA damage in firs is extremely complicated.
Alaskan forests suffered widespread defoliation, mostly by native species. The survey detected an unknown canker on quaking aspen (Populus tremuloides).
Twenty Years of Data: Trends
In addition to reporting on the 2018 survey, the more recent report contains an analysis (in Chapter 6) of data over 20 years. The analysis is intended to be used in analyses required under the Resource Protection Act (RPA). The RPA analysis uses the same set of Insect and Disease Survey data, although it groups them in four rather than five regions.
The authors grouped the annual data into four five-year windows (1996-2001; 2002-2006; 2007–2011; 2012-2016).
An overview comprising all mortality agents across the nation found a major “spurt” in area exposed to mortality-causing agents in 2002-2006. The 14.2 million ha was 4.5 times greater than the 3.1 million ha affected in the preceding 1997-2001 period. This was attributed, in part, to a sampling change. The mortality footprint fell slowly in later periods – to 9.9 million ha in 2007-2011 and 6.9 million ha in 2012-2016. Other than during the 2002-2006 period, mortality was relatively low in the Northern Region – despite EAB – especially when compared to high mortality in the Rocky Mountain and Pacific Coast Regions attributed to bark beetles.
The mortality area attributed to pathogens was also highest in 2002-2006. This was due to three pests: a root diseases-bark beetle complex killing subalpine fir, oak wilt, and beech bark disease.
Very little disease mortality was reported in the South during any of the five-year periods – an alarming failure given damage to redbay by laurel wilt. The lapse is more confusing because past IDS reports have listed redbay as heavily damaged (see USDA FS FHTET 2014).
The Pacific states saw a modest increase over time. The sudden oak death infestation was first detected by the IDS survey in 2008 – approximately 13years after it was detected on the ground.
Over the 20 years, non-native species caused mortality on an average of 500,000 ha in each five-year period except 2002-2006 (described above). The proportion of the total mortality footprint associated with non-native species was14.8% in 1997-2001, 34.4% in 2002-2006, 3.6% in 2007-2011 & 7.4% in 2012-2016.
Only in the Northern Region was a large proportion of the mortality footprint consistently attributed to non-native species – 35.1% in 1997-2011 to 98.5% in 2002-2006. The suite of species changed over time. In the first period (1997-2001), the principal species were beech bark disease, European gypsy moth, oak wilt, and hemlock woolly adelgid. During two periods – 2002-2006 and 2007-2011 – those present earlier were joined by BWA, EAB, and Dutch elm disease. By the final period – 2012-2016 – the principal agents were EAB and red pine scale.
beech bark disease Photo by Linda Haugen courtesy of Bugwood
Non-native mortality agents detected in the South included hemlock woolly agelgid and oak wilt. HWA affected 71.8% of the affected area in 2006 – 2012, 21.9% in 2012-2016. EAB was first detected by the aerial survey in 2016.
In the two western regions, non-native agents had low footprints. The highest impacts were associated with BWA and white pine blister rust in the Rocky Mountain Region, Port-Orford cedar root disease and SOD in the Pacific Region, with the addition of BWA in some years and ROD in 2015 and 2016.
Generally, woodborers have caused the highest mortality levels, seconded by pathogens. Only in the 2002-2006 period did another insect feeding guild exceed 10% of total mortality area – when BWA (a sap feeder) reached 18.7% of detected mortality – on balsam fir in Maine.
In the Western regions, the disease white pine blister rust was second to the subalpine fir mortality complex. Even this ranking fails to reflect widespread mortality of lower-elevation five-needle pines in previous decades.
In the Northern Region, a disease – beech bark disease – was most important in the first five-year period. It was replaced by a wood-boring beetle – EAB. I have noted the importance of the BWA (a sap-feeder) infestation during 2002-2006.
In the Southern Region, native bark beetles usually predominated. There were outbreaks of the foliage-feeding European gypsy moth in2007-2011 and hemlock woolly adelgid (sap-feeder) in 2012-2016.
The authors of the report note that the South might be underrepresented for several reasons – but without mentioning the severe impacts on the understory trees redbay and more recently sassafras.
SOURCES
Bailey, R.G.. 1995. Descriptions of the ecoregions of the United States. 2d ed. Miscellaneous Publication No. 1391. Washington, D.C.: U.S. Department of Agriculture Forest Service. 108 p.
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests
Guo, Q., S. Feib, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity regulates forest pest invasion. PNAS. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Morin, R.S., K.W. Gottschalk, M.E. Ostry, A.M. Liebhold. 2018. Regional patterns of declining butternut (Juglans cinerea L.) suggest site characteristics for restoration. Ecology and Evolution.2018;8:546-559
Morin, R. A. Liebhold, S. Pugh, and S. Fie. 2019. Current Status of Hosts and Future Risk of EAB Across the Range of Ash: Online Tools for Broad-Scale Impact Assessment. Presentation at the 81st Northeastern Forest Pest Council, West Chester, PA, March 14, 2019
Potter, K.M., B.S. Crane, W.W. Hargrove. 2017. A US national prioritization framework for tree species vulnerability to climate change. New Forests (2017) 48:275–300 DOI 10.1007/s11056-017-9569-5
Potter, K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019a. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests. 2019 10 304.
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019b. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation. (2019)
USDA Forest Service. Forest Health Monitoring: National Status, Trends, and Analysis 2018. General Technical Report SRS-239. June 2019. Editors Kevin M. Potter Barbara L. Conkling
USDA Forest Service. Forest Health Technology Enterprise Team. 2014. 2013-2027 National Insect and Disease Forest Risk Assessment. FHTET-14-01
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Willow forest in the Tijuana River Valley killed by Kuroshio shot hole borer the “boom” part of the cycle photo by John Boland
I have blogged earlier about the damage caused by the Kuroshio shot hole borer (KSHB, Euwallacea kuroshio), which is one of two invasive shot hole borers established in southern California. The beetle and its symbiotic fungi had caused amazing levels of damage in the Tijuana River Valley in San Diego County, California. The wood borer is described here and here.
Most of the earlier blogs focused on the absence of a response by California’s phytosanitary agency and here – until John Kabashima created sufficient political demand for a response.
A scientist who has devoted considerable effort to understanding the KSHB is John Boland of Boland Ecological Services. He has posted annual reports analyzing five years of the outbreak in the Tijuana River Valley (see full citations at the end of the blog). His principal findings (Boland and Uyeda 2020): the invasion went through a boom and bust cycle, with willows in the wettest parts of the estuary having largely recovered. So far, Kuroshio shot hole borers have not re-infested the growing trees, despite the presence of all conditions seeming to favor invasion. His principal worry is enhanced invasion by the non-native grass or reed Arundo donax.
The study site is a coastal floodplain crossed by an intermittent stream. The Tijuana River valley provides several ecosystem services, including filtering pollutants before the water reaches the ocean, open space, and important wildlife habitat, including Critical Habitat for the federally endangered least Bell’s vireo (Vireo bellii pusillus).
There is a mosaic of forests of different ages and at different distances from the current flows. They range from wet forests growing in the current river beds; dry forests growing in older river beds that get some current flows; and scrub forests growing far from current river flows. All these forests are dominated by two willows: the black willow (Salix gooddingii) and the arroyo willow (Salix lasiolepis). Both are preferred hosts of KSHB; both are pioneer species that establish in disturbed wet areas; both resprout vigorously. The riparian scrub woodlands surrounding the forests are dominated by the perennial shrub, mule fat (Baccharis salicifolia), with scattered willows of both species (Boland and Uyeda 2020).
The river carries high levels of raw sewage and industrial waste from Tijuana, Mexico. Raw sewage contains important plant nutrients – nitrogen, phosphorus and potassium. The willows in or near the nutrient-enriched channel water were growing quickly and vigorously, and had wood characteristics that differed significantly from those of trees in the dry or scrub forests. Dr. Boland notes that these trees’ phloem sap is loaded with sugars from the fast-growing leaves, and xylem sap is loaded with nutrients from the enriched soil. His Enriched Tree Hypothesis (discussed further below) suggests that these nutrients promote fast growth of the symbiotic fungi and ideal conditions for KSHB (Boland and Woodward 2019).
Boland and his colleagues have carried out detailed annual field surveys of the infestation since 2015. Using the same study plots in each year, they analyzed infestation and mortality rates, canopy damage, and survivorship of tagged willows.
Funding originally came from the U.S. Navy and U.S. Fish and Wildlife Service – agencies probably worried about the potential destruction of the vireos’ habitat. As of 2019 KSHB had infested 91% of all the willows in the valley – estimated to be more than 350,000 willows. KSHB had killed 30% of the trees, or nearly 123,000 (Boland 2019). Dr. Boland considers this estimate to be an underestimate because he could not accurately carry out surveys of individual trees in the extensively-damaged Wet Forest units in 2018. There was considerable variation in pest impacts depending on host trees’ proximity to the intermittent river. Of all of the willow deaths in the valley, 93.8% occurred in the Wet Forests, 6.1% in the Dry Forests and 0.1% in the Scrub Forests. This variation occurred even though the sites contain the same willow species (Boland and Uyeda 2020).
Infestation rates over the four-year period averaged 99% of willows in the Wet Forest units, 82% in the Dry Forests and 3% in the Scrub Woodlands. Considering 2019 alone, the overall infestation rate was only 9%. Looking at differences among forest types, 1% of the willows in Wet Forests were infested (down from 95% in 2015), 29% of the willows in dry forests (down from 73% in 2016), and 0% of the willows in the scrub forests (down from 2% in 2018). (2019 infestation rates from Boland and Uyeda 2020; earlier years from Boland 2019.)
Infestation rates had to be very high before trees died, but then mortality was very high. Only after sites reached infestation rates of more than 95% did sites have significant mortality rates – and then, very high — up to 97%. In agreement with other findings, most of the high-mortality sites were in Wet Forest units. These had a mean maximum mortality rate of 49%. The mean maximum mortality rate was only 9% in Dry Forest and 2% in Scrub Forest units (Boland and Uyeda 2020).
The size of the tree is also important. In the Wet Forests, in 2019, infestation rates were 0% for seedlings and young trees; 3% for the relatively undamaged trees that are more than 5 years old; and 1% of the resprouting adult trees that had been broken during the first wave of invasion. KSHB prefers young trees with a trunk dbh of at least 4.5 cm. Smaller trees were generally avoided. Trees with very large dbh (> 30 cm) appear to be able to survive a KSHB attack (Boland and Uyeda 2020).
At the end of the five year period, Dr. Boland had documented interesting/puzzling findings.
Recovery of the willow forest in Tijuana River Valley – 2019 photo by John Boland
Wet Forests
KSHB in the valley went through a rapid boom-and-bust cycle. In the Wet Forests, KSHB infestation progressed over the course of a few months from barely noticeable to heavy infestation and dramatic canopy collapse. Infestation rates of 80 – 95% in the West Forests in 2015 and 2016 led to virtual elimination of the canopy between 2016 and 2017 as tunnel-ridden trees were broken by wind storms. These severe damage levels occurred over 94 acres (Boland and Uyeda 2020).
After apparently depleting their preferred hosts in the wettest parts of the forest, beetle numbers fell and host trees began a rapid recovery. Mean canopy cover rose from 5% in 2017 to 56% in 2019. This recovery has taken three forms: survival of a few, scattered mature infested trees (‘Big Trees’) which grew new wood over KSHB galleries (Boland 2019); resprouting of mature KSHB-damaged trees (‘resprouts’); and seeding of new trees (‘seedlings’). Some of the forests have recovered so much in just 4 years that they are now similar to their pre-KSHB stature (Boland and Uyeda 2020).
Dr. Boland suggests that KSHB is promoted by high nutrient (pollution) levels in the water, which result in rapid growth by trees near the most steady of the intermittent streams. He has developed an Enriched Tree Hypothesis (explained briefly below; for a full discussion, see Boland and Woodland 2019).
As of autumn 2019, beetles have not attacked the recovering hosts – despite apparently favorable conditions and the absence of management interventions. 2019 infestation rates were 3% of the remaining Big Trees, 2% of the resprouting trees, 1% of the young trees, and 0% of the seedlings. Dr. Boland suggests three possible reasons (Boland and Uyeda 2020), which I will discuss below.
The resprouting trees are now old & vigorous enough to flower
It is likely that these recovering willow forests will provide good breeding habitat for least Bell’s vireo (all Boland publications).
Dry Forests
Infestation in the Dry Forests spread more slowly – infestation rates averaged 82% in the Dry Forests over four years. The infestation progressed more slowly and the canopy remained mostly intact. But in 2019 the infestation rate in the Dry Forest was substantially higher than in the Wet Forest — 29% versus 1% (Boland and Uyeda 2020).
Still, only 16% of more than 200 willows tagged in February 2016 had been killed by KSHB by autumn 2019. Among the living trees were three quarters of trees already infested in 2016, and half of trees that became infested after 2016 (Boland and Uyeda 2020).
Lack of Reinfestation (Boland and Uyeda 2020).
The absence of reinfestation is surprising, especially because the conditions thought favorable to KSHB are all present:
1. The regenerating trees belong to host species known to be preferred – black and arroyo willows.
2. The regenerating trees have reached the preferred size with trunk dbh exceeding 4.5 cm. In fall 2019 the trees in the recovering Wet Forests included many resprouting trees with mean diameters of 6.5 cm, and many new seedlings with mean diameters of 11.7 cm.
3. Recovering forests are located in the preferred nutrient-rich sites. Sewage levels remain high.
4. The trees in the recovering forests are in the condition preferred by KSHB, i.e., the trees are fast-growing and vigorous.
5. The KSHB is present – in low numbers in the Wet Forests, more numerous in Dry Forests which are < 1 km away.
It is not known whether KSHB will eventually re-infest.
The KSHB infestation reversed the presence of large trees. Originally 53% of the large trees were in the Wet Forests, 38% in the Dry Forests. The KSHB invasion damaged so many of the trees in the Wet Forests that now they represent only 24% of all the ‘Big Trees’ in the Valley; 58% of the ‘Big Trees’ are now in the Dry Forests. In many Dry Forests the remaining tall trees form a continuous canopy layer, whereas in the polluted Wet Forests they are usually only single ‘Big Trees’ (Boland and Uyeda 2020).
Other plant species
Dr. Boland expresses great concern about the spread of the invasive plant, Arundo donax, in response to canopy openings caused by the initial invasion and canopy collapse (willow trees are Arundo’s only competitors in the valley).
Surveys during fall 2019 found that most plant species growing in the Tijuana River Valley are native, dominated by the willows (mean cover of 60%). The most abundant non-native species is castor bean (10% cover). Arundo had a mean cover of only 6% in the belt transects, but it was more abundant outside the transects. Arundo is spreading as rhizomes cut loose by bulldozing, disking, and mowing on property managed by International Boundary and Water Commission (Boland and Uyeda 2020). Both willows and Arundo had increased their percent cover between 2018 and 2019 (Boland 2019).
Dr. Boland’s Recommendations (Boland and Uyeda 2020)
1) The different invasion trajectories in the three habitat types contradict some researchers’ expectation that all trajectories are similar from regardless of site characteristics or that a light infestation must be recent while a heavy infestation must be old.
2) Unique characteristics of the Tijuana River valley – especially the high sewage levels – mean that the severe infestation and damage seen there should not be expected to occur at other natural, unpolluted riparian sites.
3) Dr. Boland disputes recommendations that “heavily infested” trees be removed because they are doomed and support beetle reproduction. Although Dr. Boland studied primarily willows, he did evaluate 24 California sycamores (Platanus spp.) that had been planted in various parts of the valley.
He found that none had died and only two (8%) were infested. The two infested trees were lightly infested and growing near the sewage-enriched stream. Dr. Boland concluded that sycamores were also unlikely to be heavily infested and killed in habitats less favorable to shot hole borers (Boland 2019).
4) The ease with which native willows became densely established in wet forest sites after the first infestation wave leads Dr. Boland to advocate reliance on natural restoration projects … as long as Arundo invasion can be controlled.
5) Avoid over-fertilization or over-watering of trees in planted landscapes.
6) Focus detection searches for KSHB in nutrient-enriched areas. … e.g., near storm drain outfalls.
Research recommendations:
A) Determine why KSHB has not substantially reinvaded the recovering willow forests despite the presence of preferred species in “correct” condition and size. Dr. Boland suggests testing of three hypotheses:
Induced response of hosts. Have the infested willows changed their chemistry as a result of the borer attack, thus increasing their resistance ….
Overall forest structure. Have the less dense and more mixed forest stands reduced attractiveness to the beetle?
A disease or predator. Has a biocontrol agent been introduced accidentally? None has yet been identified …
B) Understand the possible mechanisms for the high initial infestations rates in the Wet Forests.
To evaluate the Enriched Tree Hypothesis measure the sugars and nutrients (both concentrations and loading rates) in trees subject to differing amounts of sewage or fertilizers. Then conduct controlled trials in the lab on the growth response of ISHB’s fungal symbionts to various sugar and nutrient concentrations and loading rates.
Evaluate whether willows growing in the nutrient-enriched sites produced fewer tannins that might inhibit beetle and fungal growth.
C) Can ISHB disperse by wind? Dr. Boland recommends searching for ISHB in the air high above infested trees; this could involve the use of nets or traps attached to aircraft, hot-air balloons, helium balloons or drones.
D) Determine whether surviving mature trees have superior characteristics re: morphology, vigor, and pest or disease resistance that make them less vulnerable to KSHB attack.
E) Incorporate site and ecology data and varying levels of host vulnerability into models predicting KSHB impacts. Include the ecological costs of removing the trees.
My questions
Are other scientists applying these findings in their research on KSHB or polyphagous shot hole borer outbreaks in other parts of California? I am particularly interested in the issues of possible resistance in some willows – innate or induced; and the potential role of excess nutrients in promoting fungal and beetle growth. Are they finding the ecological components of the Enriched Tree Hypothesis to be helpful in defining the impact of PSHB outbreaks in other parts of the state, and of older ages?
SOURCES
Boland, J.M. 2109. The Ecology and Management of the Kuroshio Shot Hole Borer in the Tijuana River Valley. Final Report for Naval Base Coronado under Cooperative Agreement N62473-18-2-0008
Boland, J.M. and D.L. Woodward. 2019. Impacts of the invasive shot hole borer (Euwallacea kuroshio) are linked to sewage pollution in southern Calif: the Enriched Tree Hypothesis. PeerJ 7:6812
Boland, J.M. and K.A Uyeda. 2020. The Ecology and Management of the Kuroshio Shot Hole Borer in the Tijuana River Valley 2019-20 (Year 5) Final Report. For Naval Base Coronado, Department of Navy and Southwest Wetlands Interpretive Association. Under Cooperative Agreement N62473-18-2-0008
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Another unique ecosystem being severely damaged by non-native tree-killing pests are the wetlands dominated by black ash (Fraxinus nigra). Black ash typically grows in fens, along streams, or in poorly drained areas that often are seasonally flooded. Such swamps stretch from Minnesota to Newfoundland; in the three states of Michigan, Wisconsin, and Minnesota, they cover a total of over 2 million hectares (Kolka et al. 2018).
Recent research allows us to understand the impending loss to these unique ecosystems that will be caused by the emerald ash borer (EAB).
Hydrology is the dominant factor that influences a host of ecosystem functions in black ash wetlands. Water levels are largely determined by a combination of precipitation and evapotranspiration rates. Black ash can thrive in wetter areas than most other tree species (Slesak et al. 2014). Water tables in these swamps are typically above the surface throughout early spring, followed by drawdown below the surface during the growing season with periodic rises following rain events. Water table drawdown coincides with peak evapotranspiration following black ash leaf out, demonstrating the fundamental control that this species has on animal and other plant communities (Kolka et al. 2018; Slesak et al. 2014).
Ecological Importance
Black ash generally dominate the canopy of these wetlands. Ash density can range from about 40% to almost 100%. Several other tree species are present, including northern white cedar (Thuja occidentalis), red maple (Acer rubrum), American elm (Ulmus americana) (Kolka et al. 2018), quaking aspen (Populus tremuloides), American basswood (Tilia americana), and bur oak (Quercus macrocarpa) (Slesak et al. 2014), balsam fir (Abies balsamea), balsam poplar (Populus balsamifera), and speckled alder (Alnus incana) (Youngquist et al. 2020). Black ash, by maintaining low water levels during the growing season, creates conditions under which these other trees can live but not thrive (summary of study by B.J. Palik, USDA Forest Service, here. Most other species lack the physiological adaptations of black ash or face pathogenic constraints (e.g., Dutch elm disease on American elm Ulmus americana) (Kolka et al. 2018).
Ash trees in these swamps are uneven-aged with canopy tree ages ranging from 130–232 years (Slesak et al. 2014). This complexity provides important habitat for many wildlife species, including ground beetle community assemblages (Kolka et al. 2018) and an abundance of aquatic macroinvertebrates. These are characterized and dominated by mollusks (Sphaeriidae, Lymnaeidae, Physidae), annelids (Lumbriculidae, Hirudinea), caddisflies (Limnephilidae, Leptoceridae), and dipterans (Chironomidae, Culicidae) (Youngquist et al. 2020).
a black ash swamp; source: Flickr
A major concern is that loss of trees – especially ash – might result in open marshes dominated by grasses, especially lake sedge (Carex lacustris). Conversion to sedge-dominated marshes has been observed in areas where trees have been removed as part of experiments to test various ecosystem responses to loss of the ash component (Slesak et al. 2014). Even if other trees took the place of ash, the substitutes might not support the same animal communities (see below).
Impact of Emerald ash borer and loss of black ash
Black ash is highly susceptibility to the EAB (Engelken and McCullough, 2020), so scientists expect severe impacts of the invasion in ash-dominated wetlands and – to a somewhat lesser extent — in forested stream systems’ riparian areas (Engelken and McCullough, 2020). They expect cascading impacts on 1) hydrology; 2) plant communities; 3) wildlife; 4) Native American cultures; and possibly even storage of carbon in vegetation and soils (Kolka et al. 2018).
1) Hydrology
Experiments suggest that loss of ash will cause higher water tables, especially during late summer and fall (Kolka et al 2018). This will result from reductions in evapotranspiration as large trees are replaced by shrubs and grasses (see below) (Kolka et al. 2018; Slesak et al. 2014). The higher water table might be exacerbated if higher annual precipitation levels predicted by climate change models occur. On the other hand, these models also predict a simultaneous increase in longer droughts, which might partially counteract higher precipitation and reduced evapotranspiration (Kolka et al. 2018). If they occur, these possible increases in drought length and frequency might enhance the establishment of less water-tolerant non-ash tree species in former black ash wetlands.
2) Plant Communities
Higher water tables are expected to reduce tree densities and promote conversion to open or shrub-dominated marshes. Several of the possible alternative tree species do not thrive as well as black ash under current conditions (Kolka et al. 2018). However, new hydrologic conditions might make forest restoration even more difficult because herbaceous plants transpire less water than trees, thus exacerbating the rising water tables (Slesak et al. 2014).
In upper Michigan, experiments which killed ash by cutting or girdling did not lead to an increase in growth rates of the remaining canopy species despite the increase in available resources (e.g., sunlight and nutrients) – presumably because of the raised water table (Kolka et al. 2081).
While some studies have found that black ash seedlings and saplings dominated the woody component of the swamp understory up to three years after ash were experimentally removed (Kolka et al. 2018), Engelken and McCullough (2020) found only eight saplings and a single seedling.
Scientists have planted several tree species in experiments to see which might be used to maintain the forested wetlands in the absence of black ash. The results are a confusing mix. Some species grew well once established – but had low levels of seedling establishment. Some trees planted on elevated microsites (hummocks) had the greatest survival and growth rates. (For specific data, see Kolka et al. 2018). A further consideration is tree species’ ability to adapt to warming temperatures already evident and expected to increase in coming decades (Slesak et al. 2014).
Consequently, Slesak et al. (2014) think it is likely that the EAB invasion will alter vegetation dynamics and cause a shift to an altered ecosystem state (e.g., open marsh condition) with higher water tables. They caution that the degree of ecosystem alteration will vary depending on site hydrology, annual precipitation, and period of time necessary for establishment of deeper rooted vegetation.
3) Wildlife
Moreover, any changes in vegetation will also affect the biota in more subtle ways through altered nutrient cycles. Black ash leaf litter is highly nutritious, having some of the highest nitrogen, phosphorus, and cation contents of any hardwood forest species (Kolka et al. 2018). Black ash leaves also decompose faster than most alternative tree species’ leaves (summary of Palik USDA Forest Service, here; Youngquist et al. 2018).
Youngquist et al. (2018) studied litter breakdown, litter nutritional quality, and growth of a representative invertebrate litter feeder – larvae of a shredding caddisfly (Limnephilus indivisus). They found that the larvae’s risk of death increased by a factor of three times or more when caddisflies were fed American elm, balsam poplar, or lake sedge leaves compared to black ash leaf litter. Even when the larvae lived – but matured more slowly because of the lower nutrition value of the leaves – they would still be vulnerable because they must reach metamorphosis before pond dry-down. In any planting done to maintain forested quality of wetlands, need to consider the nutritional quality of the leaf litter provided by replacements. Speckled alder was only apparently acceptable substitute; it was second to black ash in acceptability to caddisflies (Youngquist et al. 2020)
In fact, Youngquist et al. (2020) concluded that plant and detritivore biodiversity loss due to EAB invasion could alter productivity and decomposition at rates comparable to other anthropogenic stressors (e.g., climate change, nutrient pollution, acidification). The result will be altered biogeochemical cycles, resource availability, and plant and animal communities.
Scientists are also concerned about the impact of ash tree mortality on forest connectivity. Conversion of wooded swamps to shrub-and sedge-dominated wetlands will result in the loss of important micro-habitats that are already limited across the forested landscape and may also reduce availability of critical habitat for migrating birds. These changes will exacerbate on-going changes in land use in the Great Lakes region that are causing loss of forest habitat and forest homogenization. As yet, the magnitude of the impact on wildlife is unclear (Kolka et al. 2018).
black ash baskets – displayed at 2006 conference photo by Faith Campbell
4) Cultural importance – baskets
Native Americans living in the range of black ash have utilized the wood to make baskets and other tools for thousands of years. Baskets had numerous uses, such as packs for carrying items, fish traps, and for preparing food and storing household items. Ash items also had ceremonial uses and they are highly sought as gifts and in trade. The skill needed to select a good tree and work the wood is handed down through the generations and is an important part of tribes’ culture (Benedict 2010).
Discussion of these cultural traditions can be found as Powerpoints here and here.
Concerned by the spread of EAB and probable impact on black ash swamps, the USDA Forest Service has initiated major research studies with the goal of filling in the numerous knowledge gaps and developing management recommendations. A large-scale study using various manipulations to simulate the EAB invasion was initiated in the Chippewa National Forest in northern Minnesota in 2009. A companion study began in the Ottawa National Forest in Michigan in 2010 (Kolka et al. 2018). The Slesak, Youngquist, and Kolka publications cited in this blog report results of some of the studies in this project. Other studies of black ash conditions, including regeneration, at various stages of the EAB invasion wave are being carried out by Deb McCullough, Nate Siegert, and others. They are working at sites from Michigan to New England (D.G. McCullough, pers. comm.).
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report here.
For a great discussion of black ash basketweavers, see Anne Bolen, A Silent Killer: Black Ash Basket Makers are Battling a Voracious Beetle to Keep their Heritage Alive, American Indian Magazine, Spring 2020, available here.
Engelken, P.J. and D.G McCullough. 2020. Riparian Forest Conditions Along Three Northern Michigan Rivers Following Emerald Ash Borer Invasion. Canadian Journal of Forest Research. Submitted
Kolka, R.K., A.W. D’Amato, J.W. Wagenbrenner, R.A. Slesak, T.G. Pypker, M.B. Youngquist, A.R. Grinde and B.J. Palik. 2018. Review of Ecosystem Level Impacts of Emerald Ash Borer on Black Ash Wetlands: What Does the Future Hold? Forests 2018, 9, 179; doi:10.3390/f9040179 www.mdpi.com/journal/forests
Slesak, R.A., C.F. Lenhart, K.N. Brooks, A.W. D’Amato, and B.J. Palik. 2014. Water table response to harvesting and simulated emerald ash borer mortality in black ash wetlands in MN, USA. Can. J. Forestry. Res. 44:961-968.
Youngquist, M.B., C. Wiley, S.L. Eggert, A.W. D’Amato, B.J. Palik, & R.A. Slesak. 2020. Foundation Species Loss Affects Leaf Breakdown and Aquatic Invertebrate Resource Use in Black Ash Wetlands. Wetlands. Society of Wetland Scientists
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Beech trees have leafed out – so now is the time to search for symptoms of beech leaf disease.
Since its first detection near Cleveland in 2012, BLD has now been detected in 40 counties in Ohio, New York, Pennsylvania, Connecticut, and Ontario. (see map)
I ask your help now because homeowners detected the outbreaks in Connecticut in 2019. (It is often homeowners or curious citizens who detect outbreaks of tree-killing pests.)
What to look for – symptoms:
Dark bands between lateral veins of leaves. Banding is most apparent when viewing from below, looking upwards into the canopy. Banding is evident immediately upon leaf-out in the spring.
Aborted bud development and reduced leaf production.
Later stages result in heavily banded-darkened leaves that are thickened and leathery in texture, often with shriveled or curled edges.
All range of symptoms can be present on the same branch. Symptoms on individual leaves do not advance over the course of the summer. Severely affected leaves can drop off as summer progresses, sometimes as early as June. So the early season – now – is the best time to search.
Where to look? See the map of the range of American beech.
Range of American beech; source Wikimedia
In addition to checking American beech (Fagus grandifolia), also examine European beech (F. sylvatica), and Oriental beech (F. orientalis).
I encourage you to use one of the apps. However, if you are not but see something suspicious, send me a picture by using the “contact us” button. I will take a quick look, consult with experts, and – if they see what appear to be symptoms – they will tell me and I will tell you how to contact plant health authorities in your state or province.
Remember to include your email and phone number in your message to me – the “contact” form by itself does not provide sufficient information for me to respond to you.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
EAB-kiled ash tree in Shenandoah National Park in 2016 photo by F.T. Campbell
As we have known for years, forests of the eastern United States are under severe pressure from non-native forest insects and diseases. Several recent studies have put this fact into perspective.
Fei et al. (2019) found that the 15 most damaging introduced species threaten 41.1% of the total live forest biomass in the 48 conterminous states. Nine of the 15 species included in this calculation are pests of the eastern forest. Indeed, the greatest increase in biomass loss, as measured by USDA Forest Service Forest Inventory and Analysis (FIA) plot data occurred here. Compensatory growth in unaffected trees and the recruitment of new regeneration occurs only later – as much as two or more decades after the pest invasions began. Fei et al. (2019) expect these losses will be exacerbated in the future due in part to the likelihood that additional pests will be introduced.
Randall Morin found that non-native pests had caused approximately 5% increase in total mortality, by tree volume, nation-wide.
Most widespread pest threats in the East
Scientists have used several methods of measuring introduced pests’ impacts. One measure is the number of counties where the pest is present. A second measure is the proportion of the volume of the host that has been affected. Both metrics are used by Morin. A third method, used by the CAPTURE Project (Potter et al. 2019a), is the number of hosts affected by the pest.
Morin and colleagues found that the European gypsy moth has invaded 630 counties – or 29% of the volume of its principal host, oaks. (In both cases, the gypsy moth trailed white pine blister rust in extent of infestation. The latter is nationwide but having its greatest impacts in the West). The CAPTURE Project found that the gypsy moth affected the largest number of hosts – 65.
Using the “counties invaded” metric, Morin and colleagues found that dogwood anthracnose had invaded 609 counties in the East (and additional areas in the West); the emerald ash borer had invaded 479 counties at the time of analysis; the hemlock woolly adelged had invaded 432 counties. Using the number of hosts impacted measure, oak wilt (Bretziella fagacearum) affected the second largest number of hosts – 61 (Potter et al. 2019a). [All these pests are described briefly here.]
Project CAPTURE (Potter and colleagues 2019a) evaluated 339 serious pests threatening one or more of 419 native tree species in the continental US. They included both native and introduced pests. They analyzed 1,378 pest-host combinations. They found that:
54% of the host tree species (228) are infested by an exotic pest – although only 28% of the 1,378 host/agent combinations involved pests are known to be non-native in origin.
Exotic agents have, on average, considerably more severe impacts than native pests.
Non-native pests had greater average severity on angiosperms than on conifers. (As an earlier blog documented, Mech and colleagues have reached a similar – although tentative – conclusion.)
Their estimate of the threat posed by non-native pests to forests – especially for the East – is an underestimate because established pests could spread to additional vulnerable areas and there is a high likelihood that new pests will be introduced. The Southeast was consistently a “coldspot” – despite the near extirpation of one understory tree – redbay.
Potter et al. (2019a) ranked forest threats in two ways. Four host families were at highest risk to alien pests, as measured by both the numbers of tree species affected and by the most host/agent combinations: Fagaceae (oaks, tanoaks, chestnuts, beech); Pinaceae (pines); Sapindaceae (soapberry family; includes maples and buckeye); Salicaceae (willows, poplars, aspens). When host families were ranked by the severity of the host/pest threat, Fagaceae was still at greatest risk, and Sapindaceae was still in the top four; however, Ulmaceae (elms) and Oleaceae (includes Fraxinus) replaced pines and willows.
A very interesting study was published by scientists based in the Blue Ridge Mountains of Virginia (Anderson-Teixeira et al. 2020). They contend that their area is a good example of what is happening more broadly in the Mid-Atlantic region.
Anderson-Teixeira et al. (2020) found that non-native pests have substantially impacted at least 24% of the 33 tree genera (eight genera) recorded as present in their study plots. They estimated that over the century beginning with the appearance of chestnut blight in the region and ending with the expected extirpation of ash trees, net live aboveground biomass (AGB) loss among affected species totaled roughly 6.6–10 kg m -2. Forty to sixty percent of this loss started before the Park initiated quantitative surveys of permanent plots in 1987. The authors estimated that chestnut contributed up to 50% of estimated AGB losses over the century. Consequently, the estimate has very high uncertainty.
Despite these losses, Anderson-Teixeira et al. (2020) found that both total aboveground biomass and diversity within individual study plots had largely recovered through increases in non-vulnerable genera.
Average above ground biomass across the plots established in Shenandoah National Park increased as the forest recovers from logging, farming, and other disturbances before formation of the Park. These increases were due primarily to reproduction and growth of tulip poplar (Liriodendron tulipifera) and growth (but not reproduction) of oaks. Net AGB biomass was lost in oak- and hemlock-dominated plots. At plots established in the neighboring Smithsonian Conservation Biology Institute, pests had caused relatively minor impacts on AGB.
Diversity of tree species also did not change much. In the Park, the average number of genera per plot declined only 3% between 1991 and 2013. Diversity at the landscape scale increased by two genera – from 26 to 28. Many individual plots, though, lost three genera due to non-native pests – chestnut, redbud, and hemlock. A fourth genus was lost due to stochastic change. At the same time, the plots gained six native genera). This finding might be skewed by the short duration of the study period, which missed initial declines in several taxa and captured only the initial stages of decline in ash.
Several taxa were lost from the monitoring plots but were not completely extirpated from the region. Even those species not “lost” suffered elevated mortality rates and steep declines in abundance and above-ground biomass. These declines have not been reversed. The exception was some oaks, which regained above ground biomass, but not abundance, following the gypsy moth outbreak in the 1980s and early 1990s.
Fei et al. (2019) found that losses in biomass due to non-native pests – as measured by FIA plot data – was greatest for ashes, elms, beech trees, and hemlocks..
Morin and colleagues found annual mortality rates had increased three-fold above background levels for ash, beech, and hemlock. They also calculated the present mortality rates for several species for which the majority of loss occurred before their study (consequently, they could not calculate a pre-invasion “background” rate to which present rates could be compared). These included American chestnut (mortality rate of 7%), butternut (mortality rate of 5.6%), and elm trees (mortality rate of 3.5%).
The CAPTURE Project (Potter et al. 2019a) identified fifteen host-agent combinations with the highest severity. Ten of these species are found in the Mid-Atlantic region:
American chestnut (Castanea dentata)
Allegheny chinquapin (C. pumila)
Carolina ash (Fraxinus caroliniana) ,
pumpkin ash (F. profunda)
Carolina hemlock (Tsuga caroliniana)
butternut (Juglans cinerea)
eastern hemlock (Tsuga canadensis)
white ash (Fraxinus americana)
black ash (F. nigra)
green ash (F. pennsylvanica)
Four of these species are in genera included among the eight genera evaluated in the study conducted in the Blue Ridge (Anderson-Teixeira et al. 2020): American chestnut, butternut, eastern hemlock, green and white ash. The four other genera in the Blue Ridge study were elm (Ulmus), oak (Quercus), redbud Cercis, and dogwood (Cornus). All except redbud are recognized by other sources as heavily affected by non-native pests – confirming Anderson-Teixeira et al. (2020)’s conclusion that findings on the Blue Ridge reflect the wider situation.
Anderson-Teixeira et al. (2020) note that several of these tree species have been declared imperiled by the International Conservation Union (IUCN): American chestnut, butternut, American elm, eastern hemlock, and ash species.
Anderson-Teixeira et al. (2020) report data on three taxa previously important in the canopy of Blue Ridge forests – chestnut, elms, and butternut. Chestnuts larger than 10 cm DBH had disappeared from the future site of Shenandoah National Park by 1910. Short-lived sprouts continue to be present in plots in the low-elevation Smithsonian Conservation Biology Institute. Two elm species were described as ‘‘sparse’’ in the 1939 qualitative survey. Elms have persisted at low densities, low biomass, and increasingly small sizes. Butternut was ‘‘common’’ in 1939, but had disappeared from Shenandoah NP by 1987. On the Smithsonian’s property, butternut declined from four living individuals in 2008 to two in 2018. The near disappearance of butternut reflects the national picture: FIA data show the species has decreased about 58% across its U.S. range since the 1980s – which is decades after butternut canker started having a detectable impact in the Midwest.
In the Park, oak-dominated plots lost on average 24.9% of individuals and 15% of aboveground biomass. After 1995, when the gypsy moth was better controlled by spraying of Bacillus thuringiensis var. curstaki, oak aboveground biomass increased gradually, driven by individual tree growth rather than new recruitment. Continued declines in oak abundance are attributable to oak decline and management actions (or inactions) that do not promote regeneration.
In a separate study, a group of oak experts went through a process of queries to identify the greatest threat to oaks now and in the future (Conrad et al. 2020). They initially identified the following threats as most important currently (descending order): gypsy moth, oak wilt, oak decline, climate change, and drought. The top five future threats were initially identified as climate change, oak wilt, sudden oak death, oak decline, and some unknown new or emerging (non-native) pest or pathogen. By the third round, after the experts thought about their colleagues’ responses, oak decline had replaced gypsy moth as the most critical threat currently. Attack by an unknown new or emerging (non-native) pest or pathogen replaced climate change as the most critical future threat. While there was not a complete consensus, the consensus was stronger on the threat from a new pest.
remnant eastern hemlock at Linderlost, Shenandoah National Park photo by F.T. Campbell
Anderson-Teixeira et al. (2020) reported that eastern hemlock was initially present in ten of Shenandoah plots, but was no longer recorded in the survey plots after 2007. (More than 20,000 insecticide-treated trees remain alive throughout Shenandoah NP).
Before arrival of the emerald ash borer, ash aboveground biomass was increasing in Shenandoah NP and stable on the Smithsonian Institute. EAB-caused mortality was first detected at the Smithsonian site in 2016 and accelerated steeply thereafter, exceeding 12.5% year by 2018. As of 2019, ash had lost 28% of individuals and 30% of aboveground biomass relative to 2016. Ninety-five percent of remaining live trees were considered “unhealthy’’ (Anderson-Teixeira et al. 2020).
eastern (flowering) dogwood; photo by F.T. Campbell
Unlike many studies, the Shenandoah study included understory species. Flowering dogwood declined by up to 90% from plots on the Smithsonian property; 2008–2019 mortality rates averaged 7.1%. Redbud declined by up to 76% from 1995 to 2018. The 2008–2019 mortality rates averaged 6.2% year.
Anderson-Teixeira et al. (2020) concede difficulty in estimating mortality due to less virulent or lethal pathogens, including Neofusicoccum spp. on redbud and Dutch elm disease on slippery elm.
Nevertheless, they believe their analysis probably underestimates the overall pest impacts because they did not analyze several other pest/host combinations known to be present in the Park: balsam woolly adelgid (Adelges piceae) on high-elevation populations of Abies balsamea; white pine blister rust (Cronartium ribicola) on eastern white pine (Pinus strobus); beech bark disease (Neonectria spp.) on American beech (Fagus grandifolia); thousand canker disease on walnut and butternut; and emerald ash borer on the novel host fringetree Chionanthus virginicus.
Another possible threat to oaks, winter moth (Operophtera brumata), is apparently now being controlled by the biocontrol agent Cyzenis albicans.
I am uncertain about the current status of two Diplodia fungi – Diplodia corticola and D. quercivora – link to blog which have been detected in both Florida and California. In Florida, almost all the symptomatic trees grow in cultivated settings where they are exposed to various stresses (Mullerin and Smith 2015).
However, host range studies indicate that 33 species of oaks and one species of chestnut that grow in the Southeast are vulnerable, to varying degrees, to D. corticola. Oaks in the red oak group (Section Lobatae) are more vulnerable than are white oaks (Section Quercus) (Mullerin and Smith 2015). In the test, the most vulnerable appear to be the following species native to the Southeast: Q. laurifolia, Q. virginiana, Q. geminata, Q. chapmanni, Q. laevis (turkey oak), Q. phellos, Q. pumila, and Q. incana (Dreaden et al. 2016).
What should we do?
Fei et al. (2019) noted that the losses to biomass would be exacerbated by the likely introduction of additional pests. They did not recommend any prevention actions.
Conrad et al. (2020) said their findings “lend support to national regulatory and awareness efforts to prevent the introduction and establishment of novel exotic insects and pathogens.”
Anderson-Teixeira et al. (2020) join others in declaring that future survival of the IUCN-listed species probably depends on conservation and restoration actions. They cite several sources, but not the CAPTURE Project – although the two studies reinforce each other. They specifically mention limiting invasive species’ spread through strengthened regulations and “enhanced plant biosecurity cyberinfrastructure”.
This last recommendation reinforces the message of Bonello et al. (2019) link to publication. We called for creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions. One focus would be correcting the currently-inadequate focus on detection, development and deployment of genetic resistance while using modern techniques that allow for much faster breeding cycles.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
Anderson-Teixeira, K.J., V. Herrmann, W.B. Cass, A.B. Williams, S.J. Paull, E.B. Gonzalez-Akre, R. Helcoski, A.J. Tepley, N.A. Bourg, C.T. Cosma, A.E. Ferson, C. Kittle, V. Meakem, I.R. McGregor, M. N. Prestipino, M.K. Scott, A.R. Terrell, A. Alonso, F. Dallmeier, and W.J. McShea. Date? Long-Term Impacts of Invasive Insects and Pathogens on Composition, Biomass, and Diversity of Forests in Virginia’s Blue Ridge Mountains. Ecosystems
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Conrad, A.O., E.V. Crocker, X. Li, W.R. Thomas, T.O. Ochuodho, T.P. Holmes, and C. D. Nelson. 2020. Threats to Oaks in the Eastern US: Perceptions and Expectations of Experts. Journal of Forestry, 2020, 14–27
Dreaden, Black, Mullerin, and Smith. Poster presented at the 2016 USDA Invasive Species Research Forum
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. Liebhold. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National academy of Sciences.
Guo, Q., S. Feib, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity regulates forest pest invasion. PNAS. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Morin, R.S., K.W. Gottschalk, M.E. Ostry, A.M. Liebhold. 2018. Regional patterns of declining butternut (Juglans cinerea L.) suggest site characteristics for restoration. Ecology and Evolution.2018;8:546-559
Morin, R. A. Liebhold, S. Pugh, and S. Fie. 2019. Current Status of Hosts and Future Risk of EAB Across the Range of Ash: Online Tools for Broad-Scale Impact Assessment. Presentation at the 81st Northeastern Forest Pest Council, West Chester, PA, March 14, 2019
Mullerin, S. & J.A. Smith. 2015. Bot Canker of Oak in FL Caused by Diplodia corticola & D. quercivora. Emergent Pathogens on Oak and Grapevine in North America. FOR318
Potter, K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019a. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests. 2019 10 304.
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019b. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation. (2019)
Dream Lake, Rocky Mountain National Park, with limber pine photo by F.T. Campbell
In this blog, I summarize two pest threats to the unique ecosystems on high-elevation mountain ridges in the West. At risk are several keystone tree species: the five-needle pines growing at high elevations (“high-five” pines) and subalpine fir. The invasive species causing this damage – white pine blister rust (WPBR; Cronartiumribicola) and balsam woolly adelgid (BWA; Adelges piceae) – are two of the most widespread non-native species threatening North American trees and affecting the highest proportion of host volumes (Morin).
The pines being killed by white pine blister rust are whitebark pine (Pinus albicaulis), limber pine (P. flexilis), Rocky Mountain bristlecone pine (P. aristata), foxtail pine (P. balfouriana), and southwestern white pine (P. flexilis var. reflexa). As of 2010, infestations had not been reported on Great Basin bristlecone pine (P. longaeva) and the Mexican white pine species. [Unless otherwise indicated, information on white pine blister rust is from a comprehensive review and synthesis published in the August 2010 issue of Forest Pathology (Vol. 40:3-4).]
As noted above, sub-alpine fir (Abies lasiocarpa) is also being affected – although less uniformly than the pines – by the balsam woolly adelgid.
Both of these pests arrived approximately a century ago, but they are still spreading and causing additional damage. White pine blister rust had spread widely throughout the West within 40 years of its introduction. Meanwhile, BWA spread among lowland and subalpine firs along the Pacific coast from California to British Columbia within 30 years of its first detection. Its spread eastward was slower, but relentless. It reached Idaho, Montana, Utah and interior British Columbia within 50 years. Also, BWA reached Alaska within 90 years of its introduction in California. These pests are perfect examples of how invasive species introduced long ago are dreaded “gifts that keep on giving”.
For a detailed discussion of these pests’ impacts, see the descriptions posted here. To summarize, though, WPBR is present in the ranges of eight of the nine vulnerable western white pines and has caused severe mortality to some species (Sniezko et. al. 2011). For example, 88% of the limber pine range in Alberta is affected (Dawe et al. 2020). WPBR is generally causing more damage to its hosts’ northern populations. Impact of the BWA are more subtle than WPBR. Also, impacts’ severity is linked to climatic conditions. For example, measurable decline on the Olympic Peninsula was greater on south-facing slopes. However, the study did not determine whether this reflected heat-loading and tree stress or more abundant subalpine fir on these slopes. An estimated 19-53% (average 37%) of subalpine fir trees had died on sample plots on one ridge over the 19 years since BWA was first detected there. Overall forest growth after 2007 could indicate partial recovery, a momentary pause in BWA invasion, or tree growth after severe weather events (Hutton 2015).
Ranges of Trees at Risk
Many of the host trees of these two pests are widespread; others are more narrowly endemic.
Limber pine reaches from Alberta and British Columbia south to mountain peaks in Arizona and New Mexico. Whitebark pine is found from Alberta and British Columbia to California and Nevada (USDA Plants database. Subalpine fir stretches from southeast Alaska along the Canadian Rockies coast into Washington, Oregon, east into Idaho, Montana, Wyoming, Colorado, Utah, even into scattered mountain ranges of Nevada and New Mexico (Hutton 2015).
Limber pine and subalpine fir are also found in a wide range of ecosystems within these ranges. Limber pine is found at both upper and lower tree lines in grassy, open forests; on exposed rocky slopes; and in dense, mixed-conifer stands. Subalpine fir is a pioneer species on ridges, alpine meadows, avalanche chutes, and lava beds (Ragenovich and Mitchell, 2006).
Before arrival of non-native pests or pathogens, these tree species have persisted for thousands of years under harsh conditions (Hutton 2015). Many of the individual trees were long-lived; some five-needle pines, e.g., bristlecone pines, have famously live for thousands of years. Core studies demonstrated that subalpine firs trees could live 272 years in the forests of Olympic National Park and 240 years in Glacier National Park (Hutton 2015). Surely loss of these trees – or even their conversion from large and old to small and short-lived – will result in significant destruction of these unique biomes.
All these trees play important roles in high altitude, unique ecosystems (Pederson et al. no date; Dawe 2020; Hutton 2015):
They retain ground water, slow the rate of snow melt, and maintain stream flow characteristics and water quality;
They curtail soil erosion and maintain slope stability; and
They provide high-value food and shelter to wildlife.
Whitebark and limber pines are famous for providing critical food for many wildlife species at high elevations —notably bears and nutcrackers (Compendium and Dawe 2020).
More Pest Threats
Other diseases, insects, and disturbances also pose serious threats to these tree species. The threats vary by region and age of the stand. They include – for the pines — mountain pine beetle (Dendroctonus ponderosae), dwarf mistletoe (Arceuthobium spp.), and various shoot, cone or foliage insects and pathogens. For subalpine fir, threats include western balsam bark beetle (Dryocoetes confusus), fir engraver (Scolytus ventralis), and the fir root bark beetle (Pseudohylesinus granulatus) (Hutton 2015). Trees are also damaged by bear and deer, seed predation by squirrels, wildfire, and biotic succession.
On Washington’s Olympic Peninsula, BWA initiates or predisposes subalpine fir for a novel disturbance complex. BWA-caused stress makes the trees more susceptible to moisture stress and endemic bark beetle attack. Surviving trees are subsequently subject to toppling by wind. A tree can die in a few years, survive with insects for up to 20 years, or recover, depending on duration, severity, and location of infestation, and local environmental conditions (Hutton 2015).
BWA study plots in the Cascade Range experienced subalpine fir mortality ranging from 7 to 79% (measured as stem counts, not basal area) over a 19 to 38 years study period. Higher mortality occurred at low-elevation, mesic sites. One stand experienced 40% mortality in 19 years, but lost the remaining 60% during a subsequent spruce budworm infestation. Most plots continued to show sporadic signs of adelgid presence and continued tree mortality. However, 41-69% of trees survived stem infestations (Hutton 2015).
How to Protect These Ecosystems
The seeds of both whitebark and limber pines are dispersed to newly disturbed, open areas by Clark’s nutcracker (Nucifraga columbiana). Furthermore, whitebark cones open to release seeds only after fire. This had led to expectations that prescribed fire could promote regeneration of these species. However, studies by Dawe (2020) and other have found that nutcracker seed caching behavior and seedling establishment are complex. Fire management might have to vary among regions, demanding consideration of stand characteristics,like openness and the presence of other tree species. For example, in the Colorado Front Range, limber pine can be replaced by subalpine fir when fire-free intervals are long. On the other hand, in Alberta, fire appeared to boost regeneration of the dominant tree species in the stands pre-fire. In the study areas, these were white spruce (Picea glauca) and lodgepole pine (Pinus contorta) (Dawe 2020). Dawe recommends protecting existing stands of limber pine through fire mitigation efforts, e.g., thinning and other fuel treatments, and supplementary planting of seedlings.
Efforts to find biocontrol agents to target the balsam woolly adelgid began in 1957; the original focus was on the insects’ damage to Fraser fir (Abies fraseri) in the southern Appalachians. More than 25 predatory species have been introduced from Europe and Asia. There was simultaneous research on native predators. None has had an impact on BWA populations in either the East or the West.
Neither white pine blister rust nor balsam woolly adelgid is considered a quarantine pest by federal officials, so there is no attempt to prevent their movement via interstate trade in Christmas trees, timber, or nursery stock. Hutton (2015) hypothesizes that the absence of regulatory measures targetting BWA arises from the pest’s gradual effect and the hosts’ not being commercially important as timber species (although several firs are important in horticulture and as Christmas trees). I think another factor is that the pests were introduced so long ago and are now widespread.
Efforts are under way to detect resistant genotypes to be used in breeding programs. Several of the lower-elevation five-needle pines vulnerable to WPBR have benefitted from extensive breeding efforts Whitebark pine has more recently been added to programs.
The eastern Fraser fir is the target of breeding – primarily for Christmas trees (APS). However, at least small-scale volunteer efforts have been carried forward by the Alliance for Saving Threatened Forests.
Hutton (2015) expresses hope that evolutionary pressure by BWA might enhance survival of more resistant forms of subalpine fir and lead to their gradual takeover. However, I ask, why leave it to chance?
In this context, I remind you of my involvement with a group (see Bonello et al. 2019) proposing creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions – including overcoming the currently inadequate focus on detection, development and deployment of genetic resistance using modern techniques that allow for much faster breeding cycles.
I am puzzled that the Project CAPTURE places whitebark pine and subalpine fir only in Class A4.2, not among the highest priority species (Potter et al. 2019). As I blogged last spring, Project CAPTURE is part of a multi-partner effort to categorize and prioritize US tree species for conservation actions based on the threats and the trees’ ability to adapt to those threats. I find it puzzling because I am not sure I agree that these two species have a moderately high mean pest severity score – as required by the category. I am less puzzled by the assignment of a low adaptive capacity score.
Limber pine apparently ranks even lower in the Project CAPTURE priority process.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
A comprehensive review and synthesis of the history, ecology, and management of white pines threatened by white pine blister rust see the August 2010 issue of Forest Pathology (Vol. 40:3-4).
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Dawe, D.A., V.S. Peters, M.D. Flannigan. 2020. Post-fire regeneration of endangered limber pine (Pinus flexilis) at the Northern extent of its range. Forest Ecology and Management 457 (2020) 117725
Hutton, K.M. 2015. A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy. University of Washington. Available here
Morin, R. Presentation to the 81st Northeastern Forest Pest Council Northeastern states forst agencies, Philadelphia, Pennsylvania, March 2019.
Potter, K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing the conservation needs of US tree spp: Evaluating vulnerability to forest P&P threats, Global Ecology and Conservation (2019), doi: https://doi.org/10.1016/
Sniezko, R.A., M.F. Mahalovich, A.W. Schoettle, D.R. Vogler. 2011. Past and Current Investigations of the Genetic Resistance to Cronartium ribicola in High-elevation Five-needle Pines. In Keane, R.F., D.F. Tomback, M.P. Murray, and C.M Smith, eds. 2011. The future of high-elevation, five-needle white pines in Western North America. Proceedings of the High Five Symposium. 28-30 June, 2010. Missoula, MT.
The National Park Service has a legal mandate to manage lands and waters under its jurisdiction so as to “preserve unimpaired” their natural and cultural resources (NPS Organic Act 54 U.S.C. § 100101, et seq.) Invasive species undermine efforts to achieve that mission. In 2000, the NPS adopted a program to coordinate management of invasive plants. It’s not as effective as needed – see the strategic plan.
However, only recently has NPS begun trying to prioritize and coordinate programs targetting the many animals and animal diseases which threaten Park resources. These organisms range from emerald ash borer and quagga mussels; to pythons, goats, and pigs; to diseases such as white nose syndrome of bats and avian malaria in Hawai`i.
In 2017, NPS released an internal study of the pervasive threat to Park resources posed by invasive animals and discussed steps to overcome barriers to more effective responses (Redford et al., 2017; full citation at end of this blog). The Chief of the Biological Resources Division initiated this report by asking a Science Panel to evaluate the extent of the invasive animal problem, assess management needs, review best practices, and assess potential models that could serve as a service-wide organizational framework. The report was to pay particular attention to innovative and creative approaches including, but not limited to, new genomic tools. I summarized the Panel’s findings and conclusions in a blog when its report appeared in 2017.
Significantly, the
Panel’s final report states that “a general record of failure to control
invasive species across the system” was caused principally by a lack of support
for invasive species programs from NPS leadership.
This
report has now appeared in the form of a peer-reviewed article in the journal Biological Invasions by Dayer et al. 2019 (full citation at end of
this blog). Although nine of the ten authors are the same on both reports there
are substantive differences in content. For example, the journal article
reiterates the principal findings and conclusions of the Panel’s final report,
but in less blunt language.
What’s Been Watered
Down
The
toning down is seen clearly in the statements some of the panel’s six key
findings.
Finding
#1
The panel’s report says: invasive animals pose a significant threat to
the cultural and natural values and the infrastructure of U.S. national parks.
To date, the NPS has not effectively addressed the threat they pose.
Dayer et al. says: the ubiquitous presence of invasive animals in parks
undermines the NPS mission.
Finding
#2
The panel’s report says: managing
invasive animals will require action starting at the highest levels, engaging
all levels of NPS management, and will require changes in NPS culture and
capacity.
Dayer et al. says: coordinated action is required to meet the challenge.
Finding
#4
The panel’s report states: effective
management of invasive animals will require stakeholder engagement, education,
and behavior change.
Dayer et al. says: public engagement, cooperation and support is [sic]
critical.
Wording
of the other three “key findings” was also changed, but these changes are less
substantive.
Drayer
et al. also avoid the word “failure”
in describing the current status of NPS” efforts to manage invasive animal
species. Instead, these authors conclude that the invasive species threat “is
of sufficient magnitude and urgency that it would be appropriate for the NPS to
formally declare invasive animals as a service-wide priority.”
Where the
Documents Agree – Sort of
Both
the Panel’s report and Dayer et al.
state that invasive animal threats are under-prioritized and under-funded. They
say that addressing this challenge must begin at the highest levels within the
NPS, engage all levels of management, and will require investments from the NPS
leadership. Even within individual parks,
they acknowledge that staffs struggle to communicate the importance of invasive
animal control efforts to their park leadership, especially given competition with
other concerns that appear to be more urgent. And they admit that parks also
lack staff capacity in both numbers and expertise.
Also,
both the Panel’s report and Dayer et al.
urge the NPS to acknowledge formally that invasive animals represent a crisis
on par with each of the three major crises that drove Service-wide change in
the past: over-abundance of ungulates due to predator control; Yellowstone fire
crisis (which led to new wildfire awareness in the country); and recognition of
the importance of climate change.
The
Panel suggested ways to update NPS’ culture and capacity: providing incentives
for staff to (1) address long-term threats (not just “urgent” ones) and (2) put
time and effort into coordinating with potential partners, including other park
units, agencies at all levels of government, non-governmental organizations,
private landowners, and economic entities. Dayer et al. mention these barriers but does not directly mention
changing incentives as one way to overcome them.
Both
the Panel’s report and Dayer et al.
suggest integrating invasive animal threats and management into long-range
planning goals for natural and cultural landscapes and day-to-day operations of
parks and relevant technical programs (e.g., Biological Resources Division,
Water Resources Division, and Inventory and Monitoring Division).
What is Missing
from the Journal Publication
The
Panel’s final report noted the need for increased funding. It said that such
funding would need to be both consistent and sufficiently flexible to allow
parks to respond to time-sensitive management issues. It proposes several
approaches. These include incorporating some invasive species control programs
(e.g., for weeds and wood borers)
into infrastructure maintenance budgets; adopting invasive species as
fundraising challenges for non-governmental partners (e.g., “Friends of Park” and the National Park Foundation); and
adopting invasive species as a priority threat. Dayer et al. do not discuss funding issues.
The final internal report envisioned the
NPS becoming a leader on the invasive species issue by 1) testing emerging best
management practices, and 2) educating visitors on the serious threat that
invasive species pose to parks’ biodiversity. As part of this process, the
authors suggest that the NPS also take the lead in countering invasive species
denialism. Dayer et al. do not mention the issue of invasive species deniers.
Common Ground:
Status of Invasive Animals in the Parks
The
Panel’s report and Dayer et al. describe
the current situation similarly:
More than half of the National parks that responded to the internal survey (245 of the 326 parks) reported problems associated with one or more invasive animal species.
The total number of species recorded was 331. This is considered to be an underestimate since staffs often lack the ability to thoroughly survey their parks – especially for invertebrates.
Invasive species threats to Parks’ resources have been recognized for nearly 100 years. The original report notes that 155 parks reported the presence of one or more exotic vertebrate species in 1977. At that time, exotic animals were the fourth most commonly reported source of threats. In 1991, parks identified 200 unfunded projects to address exotic species, costing almost $30 million.
Only a small percentage of non-native animal invasions are under active management. Dayer et al. stated that 23% have management plans at the park unit level, and only 11% are reported as being ‘‘under control”.
Individual parks have effective programs targetting specific bioinvaders (examples are described in Redford et al; a brief summary of these efforts is provided in my previous blog.
Common Ground on
Some Solutions
The
report and Dayer et al. promote the
same steps to improve invasive animal management across the Service. Both note
that the NPS is adopting formal decision support tactics to update and
strengthen natural resource management across the board. More specific steps include
establishing
a coordination mechanism that enables ongoing and timely information sharing.
mainstreaming
invasive species issue across the NPS branches or creating a cross-cutting IAS
initiative among the Biological Resources Division, Water Resources Division,
Inventory and Monitoring Division, Climate Change Response Program, and the
regional offices.
While
both documents call on the NPS to develop and test emerging technologies, the
Panel’s final report is more detailed,
providing, in Table 5, a list of several areas of special interest, including
remotely triggered traps, species-specific toxicants, toxicant delivery
systems, drones, environmental DNA, and sterile-male releases. Dayer et al. mention eDNA and metabarcoding
for ED/RR, biocontrol, and gene drives to control invasive pathogens. (Neither
document discusses possible concerns regarding use of CRISPR and other
gene-altering technologies, other than to say there would be public concerns that
would need to be addressed.)
Both
documents note the necessity of working with resource managers beyond park
boundaries to detect and manage species before they arrive in parks. They note
that developing and operationalizing such partnerships requires time and
resources. Furthermore, invasive species prevention, eradication, and
containment programs can be effective only with public support. They suggest
strengthening NPS’ highly regarded public outreach and interpretation program
to build such support, including through the use of citizen scientists.
The
Panel’s final report said that the NPS should recognize that the condition of
the ecosystem is the objective of efforts.
Its authors recognized that achieving this goal might require
reconsidering how ecosystem management is organized within NPS so interacting
stressors (e.g., fire) and management
levers (e.g., pest eradication/suppression, prescribed fire) would be addressed.
For this, the NPS would need to create a focused capacity to address the
pressing issue of invasive animals in such a way that fosters integrated
resource management within parks, focusing on fundamental values of ecosystem
states, and not eradication targets. Dayer et
al. called for the same changes without specifically labelling “condition
of the ecosystem” as the goal.
Publication of
Dayer et al. prompted me to find out
what progress the NPS has made in responding to the “key findings” in the
Panel’s final report (neither publication calls them “recommendations”).
The
National Park Service has acted on the recommendation to appoint an “invasive
animal coordinator” within the Biological Resources Division. That person is Jennifer Sieracki. However, I wonder whether a person located in BRD is of sufficient stature to
influence agency policy across all divisions. It is not clear whether there is active
coordination with the national-level invasive plant coordinator.
Dr.
Sieriaki responded to my query by noting the following new efforts 1) to
improve outreach to partners and
the public, and 2) to expand formal and informal partnerships with local,
state, federal and tribal entities and local communities near parks.
NPS should soon finalize
two formal partnerships with other agencies and organizations for outreach and
management of invasive animal species.
NPS is working with
researchers at the US Geological Survey to expand an existing modeling tool for
identifying potential suitable habitat for invasive plant species to include
invasive insects. This will help staff focus on the most likely locations for
introductions and thus assist with early detection and control.
NPS has created a
Community of Practice so NPS employees can seek each other’s advice on addressing
invasive animal issues. A workshop of regional invasive species coordinators is
planned for the coming months to guide direction of the service-wide program
and identify other top priorities. (Seriacki pers. comm.)
I also wonder whether the NPS can achieve the top-level coordination and outreach to the public called for by both reports while complying with the terms of Public Law 116-9 – the John N. Dingle Jr. Conservation, Management, and Recreation Act, which was enacted a year ago. Title VII, Section 10(i) of this law limits spending to carry out invasive species program management and oversight to 10% of appropriated funds. Less than 15% may be spent on investigations (research), development activities, and outreach and public awareness efforts (Section 10(h)). The law does allow spending for investigations regarding methods for early detection and rapid response, prevention, control, or management; as well as inspections and interception or confiscation of invasive species to prevent in-park introductions.
For more information, see my previous criticism of NPS failure to address invasive species issues here.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
See also my earlier discussion of the new legislation here.
SOURCES
Dayer,
A.A., K.H. Redford, K.J. Campbell, C.R. Dickman, R.S. Epanchin-Niell, E.D.
Grosholz, D.E. Hallac, E.F. Leslie, L.A. Richardson, M.W. Schwartz. 2019. The
unaddressed threat of invasive animals in U.S. National Parks. Biol Invasions
Redford,
K.H., K. Campbell, A. Dayer, C. Dickman, R. Epanchin-Niell, T. Grosholz, D.
Hallac, L. Richardson, M. Schwartz. 2017. Invasive animals in U. S. National Parks:
By a science panel. Natural Resource Report NPS/NRSS/BRD/NRR—2017/1564. NPS,
Fort Collins, Colorado. Commissioned by the NPS Chief of Biological Resources
Division. https://irma.nps.gov/DataStore/DownloadFile/594922
Jennifer Sieracki, Invasive Animal Coordinator,
Biological Resources Division, National Park Service












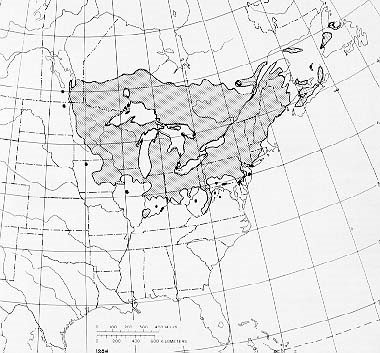


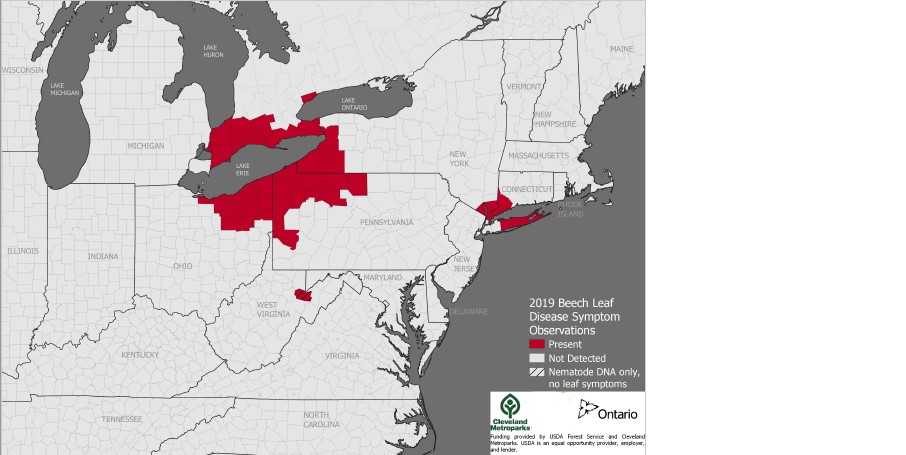
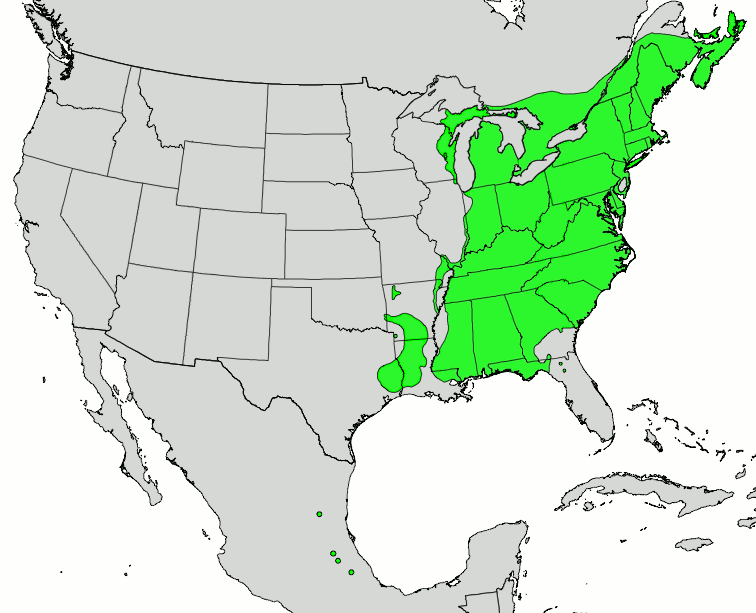





{kind=link}