
As Eastern hemlock (Tsuga canadensis) suffers high levels of mortality across nearly all its range due to hemlock woolly adelgid (HWA; Adelges tsugae), scientists scramble to determine what the successor forests will look like. The transformation will be stark: from deeply shaded evergreen coniferous forest with a sparse understory to something very different. As this process takes place, most scientists expect cascading effects on not only terrestrial and aquatic wildlife but also onecosystem functions, including soils and nutrient and hydrologic cycles (Dharmadi et al. 2019 Plotkin et al. 2024).
New England
In southern New England, hemlock groves are being replaced by stands of deciduous hardwood forests dominated by black birch (Betula lenta). While birch are expected to continue to dominate, other species comprise at least one third of seedlings in the Harvard Forest experimental sites, primarily eastern white pine (Pinus strobus) and red maple (Acer rubrum). Plotkin et al. (2024) note that conversion of hemlock forests to pine forests would be a less dramatic ecosystem shift since both are evergreen conifers.

In both southern New England and farther north, in Vermont and New Hampshire, maples and American beech have increased in prominence. In the latter case, this is despite the prevalence of beech bark disease and managers’ efforts to suppress beech. I have noted that beech leaf disease now threatens to disrupt this process.
Landowners in the region often seek to get some financial return from their forests before a pest kills the trees. About a quarter of the almost 9,000 ha of hemlock stands in the southern Connecticut River Valley have been harvested as HWA spread into the area. To test the effect of pre-mortality logging of hemlock stands, Plotkin et al. tried to mimic HWA-caused mortality by girdling all the hemlocks in some plots in Harvard Forest. In other plots they harvested most hemlocks and some of the other tree species. The girdled plots had a dramatic increase in standing and downed deadwood and a longer period of elevated understory light levels than the logged plots. They note that standing snags and on-ground dead wood provide critical ecosystem functions. Many wildlife and microbial species depend on dead wood for nutrition and a variety of micro habitats. Plotkin et al. found that the slowly decomposing dead wood also stored a large amount of carbon: girdled plots stored 18% more above-ground carbon than logged sites, even after accounting for carbon stored in harvested wood products.

The magnitude of these differences might be even larger than demonstrated in this experiment. In New England, hemlocks infested with HWA die over a decade, not the two years seen after girdling. The delayed mortality provides a longer window of opportunity for succeeding vegetation to adapt and preserve higher levels of biodiversity. Plotkin et al. (2024) suggest that logging pest-threatened hemlock forests might remove structural resources that would support forest response to ongoing climate stress and future disturbances.
Considering the disturbed plots’ invasibility by non-native plants, Plotkin et al. (2024) found that more non-native shrubs invaded the girdled plots than the logged plots. They suggest that birds that disperse the shrubs’ fleshy fruits were attracted by perch sites provided by the standing dead trees.
Southern Appalachians
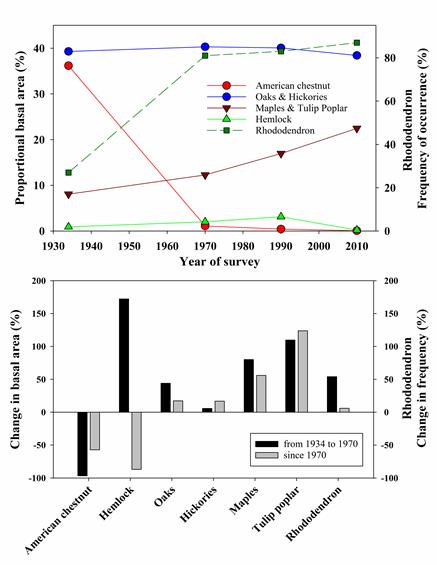
In the Southern Appalachians, post-HWA forests will apparently be quite different. At the USDA Forest Service’ Coweeta Hydrologic Laboratory in the Nantahala Mountain Range of western North Carolina, eastern hemlock died much faster than in New England. Hemlocks comprised more than 40% of the basal area before arrival of HWA (detected in 2003). Within two years all hemlock trees were infested. Half were dead by 2010, 97% by 2014 (Dharmadi et al. 2919).
In some part of the southern Appalachian forests the shrub layer is dominated by Rhododendron maximum (rosebay rhododendron). This dense shrub layer is preventing recruitment of deciduous tree species that had been expected to replace the dead hemlocks. Tree seedlings died rather than grew into saplings. Scientists working in the Coweeta experimental forest attribute the seedlings’ demise to limited access to key resources, e.g., water, nutrients (especially inorganic nitrogen), and light (Dharmadi, Elliott and Miniat 2019).
In the Coweeta Basin, hemlock loss is the most recent of a series of severe disturbances that have apparently led to a cascade of responses in the overstory, midstory, and soil that have promoted expansion of rhododendron. (The earlier disturbances were widespread logging in the 19th Century and the loss of American chestnut to chestnut blight in the first part of the 20th Century. Therefore, the response of future forests to changes in temperature and rainfall might now depend on these novel tree-shrub interactions .

R. maximum hampers succession by forming a dense subcanopy layer that greatly limits light reaching the forest floor and reduces soil moisture and temperature. These changes impede seed germination and seedling survival. In addition, rhododendron leaves that fall to the ground create a thick organic soil layer that decomposes very slowly. This affects soil chemistry, specifically availability of the key nutrient nitrogen.
The rhododendron shrubs in the region are younger than the deciduous trees now making up the canopy above them (Dharmadi, Elliott and Miniat 2019). The dense rhododendron stands resulted from the widespread mortality of American chestnut (Castanea dentata) in the early 20th century and of hemlock in the first years of the 21st Century. What’s more, even the mature deciduous trees appear to be suppressed by dense rhododendron stands. Canopy trees above rhododendrons are on average 6m shorter than those growing on sites without rhododendron thickets (Dharmadi, Elliott and Miniat 2019). In fact, by 2014, 10% of standing trees other than hemlocks had died. The tree suffering the highest level of mortality was flowering dogwood (Cornus florida). The authors do not mention a probable factor, the introduced disease dogwood anthracnose. Other species experiencing high levels of mortality are not, to my knowledge, under attack by non-native pests, so their demise seems more clearly linked to resource competition with rhododendron. These were striped maple (Acer pennsylvanicum), pitch pine (Pinus rigida), witch hazel (Hamamelis virginiana), and that staple of New England aftermath forests, black birch (Betula lenta).
Dharmadi, Elliott and Miniat (2019) suggested that managers should step in to increase recruitment in both understory and overstory layers. They proposed active management: removing rhododendrons and the soil organic layer. USFS scientists are applying these ideas experimentally at the Coweeta research station. I am unclear as to whether there is one study or more. In any case, rhododendronplants have been removed with the goal of restoring vegetation structure and composition – presumably both understory plant diversity and recruitment of tree species capable of growing into the canopy. In at least some cases, the rhododendron removal is followed by prescribed fire. One study is looking also at whether this action increased water yield.
Apparently this lack of tree regeneration is extensive – although published data are not easily accessible. Staff of the North Carolina Hemlock Restoration Initiative report they encounter similar issues (O.W. Hall, Hemlock Restoration Initiative, pers. comm.)
Several experiments have demonstrated that even in the southern Appalachians, where there are abundant moisture and rainfall, the trees and shrubs compete for water and other nutrients. However, Dharmadi et al. (2022) found that removal of the rhododendron shrub layer is unlikely to significantly alter streamflow, atr least during the growing season. In winter, when deciduous trees lack leaves, reduction in interception of precipitation might result in increased streamflow (Dharmadi et al. 2022). I ask whether increasing stream flow in winter is a goal? I thought the concern was stream flow levels in summer.
Nor is removal of the rhododendron shrub layer likely to alter stream chemistry during the growing season.
Removal of living Rhododendron and leaf litter apparently can help restore forest structure through improving tree seedling survival and recruitment as well as increasing growth of established trees.
Removing Privet
However, other management actions might bring about desired changes more effectively or broadly. Specifically Dharmadi and colleagues mentioned removal of privet (Ligustrum) – a very widespread invasive shrub in forests of the Southeast. (Fifteen years ago it was estimated that just one privet species, Chinese privet, occupied more than a million hectares in 12 southeastern states [Hanula 2009].)

I ask also whether prescribed fire to remove the rhododendron-dominated soil organic layer is useful. Dharmadi and colleagues found that such fires reduced leaf litter temporarily, but annual leaf-fall replaced the litter layer the next year, so this management effort is unlikely to affect plot evapotranspiration rates.
Supporting Pollinators
Another study (Ulyshenet al. 2022) examined whether removing rosebay rhododendron would benefit bees and other pollinators. They found that removal of Rhododendron alone (without fire) did not dramatically improve pollinator habitat in the southern Appalachians. In fact, about a quarter of the bee species studied visited R. maximum flowers and might decline if the shrub’s population is reduced. Ulyshen and colleagues suggest that some factors that correlate with fire severity probably promotes growth of insect-pollinated plants. They suggest specifically the greater presence of downed woody debris, which provides nesting sites and other resources used by insects. They recommended creation of open areas to support wildflowers as a more effective way to benefit bees in this region. Again, rhododendron removal pales in effectiveness compared to eradication of privet.
SOURCES
Dharmadi, S.N., K.J. Elliott, C.F. Miniat. 2019. Lack of forest tree seedling recruitment and enhanced tree and shrub growth characterizes post-Tsuga canadensis mortality forests in the southern Appalachians. Forest Ecology and Management 440 (2019) 122–130.
Dharmadi, S.N., K.J. Elliott, C.F. Miniat. 2022. Larger hardwood trees benefit from removing Rhododendron maximum following Tsuga canadensis mortality. Forest Ecology and Management
Hanula, J.L., S. Horn, and J.W. Taylor. 2009. Chinese Privet (Ligustrum sinense) Removal and its Effect on Native Plant Communities of Riparian Forests. Invasive Plant Science and Management 2009 2:292–300.
Plotkin, A.B., A.M. Ellison, D.A. Orwig, M.G. MacLean. 2024. Logging response alters trajectories of reorganization after loss of a foundation tree species. Ecological Applications. 2024;e2957.
Ulyshen, M., K. Elliott, J. Scott, S. Horn, P. Clinton, N. Liu, C.F. Miniat, P. Caldwell, C. Oishi, J. Knoepp, P. Bolstad. 2022. Effects of Rhododendron removal and prescribed fire on bees and plants in the southern Appalachians. Ecology and Evolution. 2022;12:e8677.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
or







































{kind=link}