American bullfrog (Lithobates catesbeianus); photo by Will Brown via Wikimedia; one of invasive animals deliberately introduced to Europe in the past
In February 2024 the European Parliament approved legislation outlawing “ecocide” and providing sanctions for environmental crimes. Member states now have two years to enshrine its provisions in national law.
The new rules update the list of environmental crimes adopted in 2008 and enhance the sanctions. The goal is to ensure more effective enforcement. Listed among the offenses are:
the import and use of mercury and fluorinated greenhouse gases,
the import of invasive species,
the illegal depletion of water resources, and
pollution caused by ships.
This action followed an in-depth analysis of the failures of the previous EU environmental directive, first adopted in 2008 (Directive 2008/99/EC). The review found that:
The Directive had little effect on the ground.
Over the 10 years since its adoption few environmental crime cases were successfully investigated and sentenced.
Sanction levels were too low to dissuade violations.
There had been little systematic cross-border cooperation.
EU Member states were not enforcing the Directive’s provisions. They had provided insufficient resources to the task. They had not developed the needed specialized knowledge and public awareness. They were not sharing information or coordinating either among individual governments’ several agencies or with neighboring countries.
The review found that poor data hampered attempts by both the EU body and national policy-makers to evaluate the Directive’s efficacy.
The new Directive attempts to address these weaknesses. To me, the most important change is that complying with a permit no longer frees a company or its leadership from criminal liability. These individuals now have a “duty of care”. According to Antonius Manders, Dutch MEP from the Group of the European People’s Party (Christian Democrats), if new information shows that actions conducted under the permit are “causing irreversible damage to health and nature – you will have to stop.” This action reverses the previous EU environmental crime directive – and most member state laws. Until now, environmental crime could be punished only if it is unlawful; as long as an enterprise was complying with a permit, its actions would not be considered unlawful. Michael Faure, a professor of comparative and international environmental law at Maastricht University, calls this change revolutionary.
Lorton Prison; via Flickr
Another step was to make corporate leadership personally liable to penalties, including imprisonment. If a company’s actions cause substantial environmental harm, the CEOs and board members can face prison sentences of up to eight years. If the environmental harm results in the death of any person, the penalty can be increased to ten years.
Financial penalties were also raised. Each Member state sets the fines within certain parameters. Fines may be based on either a proportion of annual worldwide turnover (3 to 5%) or set at a fixed fine (up to 40 million euros). Companies might also be obliged to reinstate the damaged environment or compensate for the damage caused.Companies might also lose their licenses or access to public funding, or even be forced to close.
Proponents of making ecocide the fifth international crime at the International Criminal Court argue that the updated directive effectively criminalizes “ecocide”— defined as “unlawful or wanton acts committed with knowledge that there is a substantial likelihood of severe and either widespread or long-term damage to the environment being caused by those acts.”
Individual member states also decide whether the directive will apply to offences committed outside EU borders by EU companies.
Some members of the European Parliament advocate for an even stronger stance: creation of a public prosecutor at the European Union level. They hope that the Council of Europe will incorporate this idea during its ongoing revision of the Convention on the Protection of the Environment through Criminal Law. To me, this seems unlikely since the current text of the Convention, adopted by the Council in 1998, has never been ratified so it has not come into force.
The Council of Europe covers a wider geographic area than the European Union – 46 member states compared to 27. Members of the Council of Europe which are not in the EU include the United Kingdom, Norway, Switzerland, Bosnia-Hercegovina, Serbia, Kosovo, Albania; several mini-states, e.g., Monaco and San Remo; and countries in arguably neighboring regions, e.g., Armenia, Azerbaijan, Georgia, and Turkey.
While I rejoice that invasive species are included in the new Directive, I confess that I am uncertain about the extent to which this inclusion will advance efforts to prevent spread. The species under consideration would apparently have to be identified by some European body as “invasive” and its importation restricted. As we know, many of the most damaging species are not recognized as invasive before their introduction to a naïve environment. On the other side, the requirement that companies recognize new information and halt damaging actions – even when complying with a permit! – provides for needed flexibility.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
U.S. Department of Agriculture headquarters; lets lobby these people! photo by Wikimedia
Twenty-three scientists based around the world published a Letter to the Editor titled “Overwhelming evidence galvanizes a global consensus on the need for action against Invasive Alien Species” It appears in the most recent edition of Biological Invasions (2024) 26:621–626.
The authors’ purpose is to draw attention to the release of a new assessment by the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services’ (IPBES).
The report was issued in September 2023. It is described as the most comprehensive global synthesis of the current knowledge on the bioinvasion process and the impacts of invasive alien species (952 pages!). Its preparation took nearly a decade. Most important, it represents the first consensus among governments and scientists worldwide on the magnitude and extent of the threats that bioinvasions pose to nature, people, and the economy.
The proposed solutions are astoundingly broad and ambitious: transformation of how governments and societies perform. I don’t disagree! However, we need interim steps – “bites of the elephant.” In my view, the report falls short on providing these.
Our challenge: join others in bringing this analysis to decision-makers’ attention. Can we pull out information that will help persuade U.S. decision-makers – governmental and non-governmental – that the threat is both urgent and solvable? How do we more effectively advocate for the aggressive, science-based action that we all know is needed?
(I hope that the fact that the report was prepared under the auspices of the Convention on Biodiversity, to which the U.S. is not a party, does not intensify the challenge for us.)
Why we need to restructure the behavior of governments and societies
Bioinvasions are facilitated by policies, decision-making structures, institutions, and technologies that are almost always focused on achieving other goals. Species transport and introduction are driven by policies aimed at promoting economic growth – especially trade. Later stages of invasions, i.e., establishment and some spread, are accelerated by certain uses of land and sea plus climate change. For example, activities that fragment habitats or cause widespread habitat disturbance provide ready places for bioinvasions. Rarely are those who gain by such policies held accountable for the harms they produce via bioinvasions.
To address these unintended consequences, the IPBES report calls for “integrated governance.” Its authors want coordination of all policies and agencies that touch on the indirect drivers, e.g., conservation; trade; economic development; transport; and human, animal, and plant health. Policy instruments need to reinforce – rather than conflict with — strategic invasive species management across sectors and scales. This involves international agreements, national regulations, all governmental sectors, as well as industry, the scientific community, and ordinary people – including local communities and Indigenous Peoples.
The report also calls for establishment of open and inter-operable information systems. This improved access to information is critical for setting priorities; evaluating and improving regulations’ effectiveness; and reducing costs by avoiding duplication of efforts.
Critically important information that is often unspoken:
Indirect causes underlying the usual list of human activities that directly promote bioinvasions are the rapid rise of human population and even more rapid rise in consumption and global trade.
Biosecurity measures at international borders have not kept pace with the growing volume, diversity, and geographic origins of goods in trade.
Continuation of current patterns is expected to result in one-third more invasive species globally by 2050. However, this is an underestimate because today’s harms reflect the consequences of past actions – often from decades ago. Drivers of invasions are expected to grow in both volume and impact.
We can prevent and control invasive alien species – but that success depends on the availability of adequate, sustained resources, plus capacity building; scientific cooperation and transfer of technology; appropriate biosecurity legislation and enforcement; and engaging the full range of stakeholders. These require political will.
A major impact of bioinvasion is increased biotic homogenization (loss of biological communities’ uniqueness). This concerns us because we are losing the biotic heterogeneity that provides insurance for the maintenance of ecosystem functioning in the face of ongoing global change.
The IPBES study asserts that successfully addressing bioinvasions can also strengthen the effectiveness of policies designed to respond to other drivers, especially programs addressing conservation of biological diversity, ensuring food security, sustaining economic growth, and slowing climate change. All these challenges interact. The authors affirm that evidence-based policy planning can reflect the interconnectedness of the drivers so that efforts to solve one problem do not exacerbate the magnitude of others and might even have multiple benefits.
More Key Findings
Overall, 9% (3,500) of an estimated 37,000 alien species established in novel environments are invasive (those for which scientists have evidence of negative impacts). Proportions of invasives is high among many taxonomic groups: 22% of all 1,852 alien invertebrates; 14% of all 461 alien vertebrates; 11% of all 141 alien microbes; and 6% of all 1,061 alien plants. (The discussion of probable undercounts relates to aquatic systems and certain geographic regions. However, I believe these data are all undermined by gaps in studies.)
Invasive alien species – solely or in combination with other drivers – have contributed to 60% of recorded global extinctions. Invasive species are the only driver in 16% of global animal and plant extinctions. Some invasive species have broader impacts, affecting not just individual species but also communities or whole ecosystems. Sometimes these create complexoutcomes that push the system across a threshold beyond which ecosystem restoration is not possible. (No tree pests are listed among the examples.)
dead whitebark pine in Glacier National Park; photo by National Park Service
The benefits that some non-native – even invasive – species provide to some groups of people do not mitigate or undo their negative impacts broadly, including to the global commons. The report authors note that beneficiaries usually differ from those people or sectors that bear the costs. The authors cite many resulting inequities.
There are insufficient studies of, or data from, aquatic systems, and from Africa; Latin America and the Caribbean; and parts of Asia.
The number of alien species is rising globally at unprecedented and increasing rates. There are insufficient data specifically on invasive species, but they, too, are thought to be rising at similar rates.
Horticulure is a major pathway for introducing 46% of invasive alien plant species worldwide.
Regarding invasive species’ greater impact on islands,the IPBES report mentions brown tree snakes on Guam and black rats on the Galapagos Islands. It also notes that on more than a quarter of the world’s islands, the number of alien plants exceeds the total number of native ones. See my blogs on non-native plants on Hawai`i and Puerto Rico. In addition, I have posted several blogs regarding disease threats to rare bird species in Hawai`. The IPBES report does not mention these.
Where the Report Is Weak: Interim Steps
The report endorses adoption of regulated species (“black”) lists.
The report emphasizes risk analysis of species. Unfortunately IPBES’ analysis was completed before publication of the critique of risk analysis methods by Raffa et al. ( (2023) (see references). However, we must take the latter into consideration when deciding what to advocate as U.S. policy.
The report authors call for more countries to adopt national legislation or regulations specifically on preventing and controlling invasive species. (They note that 83% of countries lack such policies). They also list the many international agreements that touch on invasive species-relevant issues. However, Raffa et al. found that the number of such agreements to which a country is a party bears no relationship to the numbers of alien species detected at its border or established on its territory.
The challenge to risk assessment posed by multiple sources of uncertainty can be managed by recognizing, quantifying, and documenting the extent of that uncertainty.
Beech leaf disease – one of many non-native pests that were unknown before introduction to a naive ecosystem. Photo by Jennifer Koch, USDA Forest Service
I appreciate the report’s emphasis on the importance of public awareness and engagement, but I thought the discussion of effective campaigns lacked original ideas.
The report did not fulfill its own goal of fully exploring unappreciated impacts of policies in its discussion of habitat fragmentation. For example, the report notes that grazing by feral alien ungulates facilitates the spread of invasive alien plant species. However, it does not mention the similar impact by livestock grazing (Molvar, et al. 2024).
SOURCES
Molvar, E.M., R. Rosentreter, D. Mansfield, and G.M. Anderson. 2024. Cheat invasions: History, causes, consequences, and solutions. Hailey, Idaho: Western Watersheds Project, 128 pp.
Raffa, K.F., E.G. Brockerhoff, J-C. GRÉGOIRE, R.C. Hamelin, A.M. Liebhold, A. Santini, R.C. Venette, and M.J. Wingfield. 2023. Approaches to forecasting damage by invasive forest insects and pathogens: a cross-assessment. BioScience 85 Vol. 73 No. 2 (February 2023) https://academic.oup.com/bioscience
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Clive Braser and others study Phytophthora species in their native habitats of Vietnam; which will become aggressive invaders in North America?
For years, one focus of this blog has been on scientists’ efforts to improve prevention of new introductions of forest pests. In earlier blogs, I summarized and commented on efforts by Mechet al. (2019) and Schultzet al. (2021), who extrapolate from insect-host relationships of pests already established in North America. [Full citations are presented at the end of this blog.] Both limited their analysis to insects; Mech et al. focused on those that attack conifers, Schultz et al. on those that attack single genera of angiosperms (hardwoods).
However, many of the most damaging agents are pathogens; for an indication, review the list under “invasive species” here. Indeed, Beckman et al. (2021) reported that only three non-native organisms pose serious threats to one or more of the 37 species of Pinus native to the U.S. All are pathogens: white pine blister rust (WPBR), pitch canker, and Phytophthora root rot (Phytophthora cinnamomi).
For this reason I welcome a study by Li et al. (2023), who used laboratory tests to evaluate the threat posed by more than 100 fungi associated with bark beetles. Since there are more than 6,000 species of bark and ambrosia beetles and they are commonly intercepted at the U.S. border, determining which should be priorities is important. Li et al. point out that the vast majority of such introductions have had minimal impacts. Two, however, have caused disastrous levels of damage: Dutch elm disease and laurel wilt disease.
Li et al. tested 111 fungi associated with 55 scolytine beetles from areas of Eurasia with latitudes and ecosystems analagous to those in the southeastern U.S. The beetles assessed included beetle species responsible for recent major tree mortality events in Eurasia: Dendroctonus species, Platypus koryoensis (Korean oak wilt), Platypus quercivorus (Japanese oak wilt) and Tomicus species.
The authors tested the fungi’s virulence on four species of trees native to the Southeast – two pines (Pinus taeda and P. elliottii var. elliottii), and two oaks(Quercus shumardii and Q. virginiana).
Li et al. found that none of 111 fungal associates caused a level of damage on these four hosts equal to Dutch elm disease on elms or laurel wilt disease on trees in the Lauraceae. Twenty-two of the fungi were minor pathogens – meaning they might cause damage under certain conditions or when loads of inoculum are large enough.
redbay trees killed in coastal Georgia by laurel wilt; photo by Scott Cameron
I think Li et al. set an extremely high bar for “serious” damage. Surely we wish to prevent introduction of pathogens that cause damage at a lower level than the catastrophes to which these two diseases have exposed a genus (elms) and a family (Lauraceae)! Still, the scientific approach used here is a step toward addressing pathogens. These agents of tree mortality are addressed much less frequently than insects. I hope that scientists will continue to test the virulence of these fungi on some of the thousands of other species that make up the forests of the United States, or at least the dominant species in each ecosystem.
It is discouraging that Raffaet al. (2023) found none of four approaches to predicting a new pest’s impact to be adequate by itself. Instead, they outlined the relative strengths and weaknesses of each approach and the circumstances in which they might offer useful information. I am particularly glad that they have included pathogens, not just insects. The four approaches they review are:
(1) pest status of the organism in its native or previously invaded regions;
(2) statistical patterns of traits and gene sequences associated with high-impact pests;
(3) sentinel plantings to expose trees to novel pests; and
(4) laboratory tests of detached plant parts or seedlings under controlled conditions.
They emphasize that too little information exists regarding pathogens to predict which microbes will become damaging pathogens when introduced to naïve hosts in new ecosystems. See the article, especially Figure 4, for their assessment of the strengths each of the several approaches.
Raffa et al. raise important questions about both the science and equity issues surrounding invasive species. As regards scientific issues, they ask, first, whether it will ever be possible to predict how each unique biotic system will respond to introduction of a new species. Second, they ask how assessors should interpret negative data? In the context of equity and political power, they ask who should make decisions about whether to act?
In my blog I expressed concern about finding that most introduced forest insects are first detected in urban areas whereas introduced pathogens are more commonly detected in forests. I hope scientists will redouble efforts to improve methods for earlier detection of pathogens. Enrico Bonello at Ohio State and others report that spectral-based tools can detect pathogen-infected plants, including trees.
Japanese cherry trees burned on the Washington D.C. mall because infested by scale; on order of Charles Marlatt
International trade is considered the single most important pathway for unintentional introductions of insects. Updated figures remind us about the stupendous amounts of goods being moved internationally. According to Weber et al., international shipping moves ~133 million TEU containers per year between countries, the majority between continents. Four times this number move within regions via coastal shipping. On top of that, four billion passenger trips take place by air every year. Air freight carries another ~220 million tons of goods; while this is a tiny fraction of the weight shipped by boat, the packages are delivered in less than a day – greatly increasing the likelihood that any unwanted living organisms will survive the trip. The U.S. also imports large numbers of live plants – although getting accurate numbers is a challenge. MacLachlan et al. (2022) report 5 billion plants imported in 2021, but the USDA APHIS annual report for FY22 puts the number at less than half that figure: 2.2 billion plant units.
Given the high volume of incoming goods, Weber et al. advocate improved surveillance (including analysis of corresponding interceptions) of those pathways that are particularly likely to result in non-native species’ invasions, e.g. live plants, raw lumber(including wood packaging), and bulk commodities e.g. quarried rock. Isitt et al. and Fenn-Moltu et al. concur that investigators should focus on the trade volumes of goods that are likely to transport plant pests – in their cases, plant imports.
The importance of the plant trade as a pathway of introduction for has been understood for at least a century – as witnessed by the introductions of chestnut blight DMF and white pine blister rust, DMF and articles by Charles Marlatt. A decade ago, Liebhold et al. (2012) calculated that the approach rate of pests on imported plants was 12% — more than 100 times higher than the 0.1% approach rate found by Haack et al. (2014) for wood packaging.
Since plant-insect interactions are the foundation of food webs, changes to a region’s flora will have repercussions throughout ecosystems, including insect fauna. See findings by teams led by Doug Tallamy and Sara Lalk; and a chapter in the new forest entomology text written by Bohlmann, and Krokene (citation at end of blog under Allison, Paine, Slippers, and Wingfield). Sandy Liebhold and Aymeric Bonnamour also addressed explicitly links between introductions of non-native plant and insect species. Weber et al. call this phenomenon the “receptive bridgehead effect”: a non-native plant growing prolifically in a new ecosystem provides a suitable host for an organism that feeds on that host, raising the chance for its establishment.
Recent studies confirm the importance of the “receptive bridgehead effect”. Isitt and colleagues found that the large numbers of introduced European insect species – all taxa, not just phytophagous insects – established in North America and Australia/New Zealand were best explained by the numbers of European plants introduced to these regions – in other words, the most important driver appears to be the diversity of non-native plants.
The presence of European plants in North America and Australia/New Zealand promoted establishment of European insects in two ways. First, these high-volume imports increased the propagule pressure of insects associated with this trade. Live plant imports might have facilitated the establishment of ~70% of damaging non-native forest insects in North America. Second, naturalization of introduced European plants provided a landscape replete with suitable hosts. This is especially obvious in Australia/New Zealand, which have unique floras. In Australia, nearly 90% of non-native pest insects are associated with non-native plants. Those non-native insects that do feed on native plants are more likely to be polyphagous.
Amur honeysuckle – one of the hundreds of Asian plants invading North American ecosystems; via Flickr
I hope U.S. phytosanitary officials apply these lessons. Temperate Asia is the source of more non-native plants established in both North America and Australia/New Zealand than is Europe. Already, many insects from Asia have invaded the U.S. The logicof the “receptive bridgehead effect” points to prioritizing efforts to prevent even more Asian insects from reaching our shores!
Fenn-Moltu et al. sought to elucidate which mechanisms facilitate species’ success during the transport and introduction/establishment stages of bioinvasion. They studied the transport stage by analyzing border interceptions of insects from 227 countries by Canada, mainland U.S., Hawai`i, Japan, New Zealand, Great Britain, and South Africa over the 60 year period 1960 – 2019. They studied establishment by analyzing attributes of 2,076 insect species recorded as established after 1960 in the above areas plus Australia (North America was treated as a single unit comprised of the continental U.S. and Canada).
The number of species transported increased with higher Gross National Income in the source country. The number of species transported decreased with geographic distance. They suggest that fewer insects survive longer journeys, but say additional information is needed to verify this as the cause. The number of species transported was not affected by species richness in the native region.
More species established when introduced to a country in the same biogeographic region. They were not surprised that environmental similarity between source and destination apparently strongly affected establishment success. The number of species established was not affected by species richness in the native region. For example, the greatest number of established species originated from the Western and Eastern Palearctic regions, which together comprise only the fifth-largest pool of native insect species.
Gaps Despite Above Studies
As I noted at the beginning, most of the studies examining current levels of pests transported on imported plants have been limited to insects. This is unfortunate given the impact of introduced pathogens (again, review the list damaging organisms under “invasive species” here).
In addition, most studies analyzing the pest risk associated with plant imports use port inspection data – which are not reliable indicators of the pest approach rate. The unsuitability of port inspection data was explained by Liebhold et al. in 2012 and Fenn-Moltu et al. a decade later – as well as Haack et al. 2014 (as the data pertain to wood packaging). Fenn-Moltu et al. note that inspection agencies often (and rightly!) target high-risk sources/commodities, so the records are biased. Other problems might arise from differences in import volume, production practices, and differences in records that identify organism only to genus level rather than species. Fenn-Moltu et al. call for relying on randomized, statistically sound inspection systems; one such example is USDA’s Agriculture Quarantine Inspection System (AQIM). Under AQIM, incoming shipments are randomly selected and put through more thorough inspections to produce statistically based estimates of approach rates, defined as the percent of inspected shipments found to be infested with potential pests (Liebhold et al. 2012). I ask why scientists who are aware of this issue have not obtained AQIM data for pests associated with plant imports. Plant imports have been included in the AQIM system since 2008. Have they not been able to persuade APHIS to provide these data? Or are these data available for only limited types of imported plants? Too narrow a focus would create a different source of potential bias.
Both Isitt et al. and Fenn-Moltu et al. list factors not addressed and other caveats of which we should be aware when extrapolating from their findings.
Beckman, E., Meyer, A., Pivorunas, D., Hoban, S., & Westwood, M. (2021). Conservation Gap Analysis of Native U.S. Pines. Lisle, IL: The Morton Arboretum.
Fenn-Moltu, G., S. Ollier, O.K. Bates, A.M. Liebhold, H.F. Nahrung, D.S. Pureswaran, T. Yamanaka, C. Bertelsmeier. 2023. Global flows of insect transport and establishment: The role of biogeography, trade and regulations. Diversity and Distributions DOI: 10.1111/ddi.13772
Hoddle. M.S. 2023. A new paradigm: proactive biological control of invasive insect pests. BioControl https://doi.org/10.1007/s10526-023-10206-5
Isitt, R., A.M. Liebhold, R.M. Turner, A. Battisti, C. Bertelsmeier, R. Blake, E.G. Brockerhoff, S.B. Heard, P. Krokene, B. Økland, H. Nahrung, D. Rassati, A. Roques, T. Yamanaka, D.S. Pureswaran. 2023. Drivers of asymmetrical insect invasions between three world regions. bioRxiv preprint doi: https://doi.org/q0.1101/2023.01.13.523858
Li, Y., C. Bateman, J. Skelton, B. Wang, A. Black, Y-T Huang, A. Gonzalez, M.A. Jusino, Z.J. Nolen, S. Freemen, Z. Mendel, C-Y Chen, H-F Li, M. Kolarik, M. Knizek, J-H. Park, W. Sittichaya, T-H Pham, S. Ito, M. Torii, L. Gao, A.J. Johnson, M. Lu, J. Sun, Z. Zhang, D.C. Adams, J. Hulcr. 2022. Pre-invasion assessment of exotic bark beetle-vectored fungi to detect tree-killing pathogens. Phytopathology Vol 112 No. 2 February 2022
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Liebhold, A.M., T. Yamanaka, A. Roques, S. August, S.L. Chown, E.G. Brockerhoff and P. Pyšek. 2018. Plant diversity drives global patterns of insect invasions. Sci Rep8, 12095 (2018). https://doi.org/10.1038/s41598-018-30605-4
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Mech, A.M., K.A. Thomas, T.D. Marsico, D.A. Herms, C.R. Allen, M.P. Ayres, K.J. K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, and P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol. 2019 Nov; 9(21): 12216–12230.
Raffa, K.F., E.G. Brockerhoff, J-C. Gregoirem R.C. Hamelin, A.M. Liebhold, A. Santini, R.C. Venette, and M.J. Wingfield. 2023. Approaches to Forecasting Damage by Invasive Forest Insects and Pathogens: A Cross-Assessment. Bioscience Vol. 73, No. 2. February 2023.
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions.
Weber, D.C., A.E. Hajek, K.A. Hoelmer, U. Schaffner, P.G. Mason, R. Stouthamer, E.J. Talamas, M. Buffington, M.S. Hoddle and T. Haye. 2020. Unintentional Biological Control. Chapter for USDA Agriculture ResearchService. Invasive Insect biocontrol and Behavior Laboratory. https://www.ars.usda.gov/research/publications/?seqNo115=362852
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
I have blogged many times about the risk of pest introductions on imports of live plants [= “plants for planting” in USDA’s terms]. Last October I reviewed 14-year old data indicating that nearly 70% of 455 damaging tree pests introduced to the continental U.S. had probably been introduced via plant imports. These included 95% of sap feeding and 89% of foliage feeding insects and about half of the pathogens. The approach rate of pests on imported plants was apparently 12% (Liebhold et al. 2012) — more than 100 times higher than the 0.1% approach rate found by Haack et al. (2014) for wood packaging.
First, those analyses focus almost exclusively on insects (MacLachlan et al. 2022 focused on a single insect order, the Hemiptera!), despite the many pathogens probably introduced by the plant trade in recent decades. Examples I cited included several Phytophthoras, rapid ohia death, beech leaf disease, and boxwood blight. There have been repeated detections of the Ralstonia solanacearum Race 3 biovar 2.
SOD- infected rhododendrons; photo by Jennifer Parke, Oregon State University
Second, most studies analyzing the pest risk associated with plant imports use port inspection data – which are not reliable indicators of the pest approach rate – as explained by Liebhold et al. 2012 and Haack et al. 2014 (as it pertains to wood packaging).
Third, many of the studies are based on data from a decade or longer in the past. This means the studies do not address whether APHIS’ recent changes in its approach – including adoption of NAPPRA – have resulted in reduced introductions.
A complication is that, since insects are difficult to detect, those associated with the high volumes of plants imported in recent years might not be detected for years or decades after their introduction.
I have called for APHIS to update the Liebhold et al. 2012 study to determine the approach rate for all types of organisms that threaten North American tree species. Any such study should include trees on Hawai`i, Guam, Puerto Rico, and other U.S possessions and territories. These islands are nearly always excluded from analyses of imported pests. I concede that there are probably scientific and data-management challenges but these islands are immensely important from a biodiversity point of view, and they are parts of the United States!
eastern hemlocks killed by hemlock woolly adelgid; Linville Gorge; photo by Steven Norman, USFS
MacLachlan et al. (2022) estimated that new establishments – of insects in the order Hemiptera – per unit of additional plant imports have shrunk substantially. They attribute this decline to a combination of increased imports and the presence of a growing number of insect species introduced in the past. They found that introductions to the Asian Palearctic and Neotropic regions have been reduced by depletion of species pools. Other factors are thought to explain the substantial decline in establishment likelihood for the other regions. However, lag times in detecting insect introductions complicate this assessment.
However, despite that significant decrease in risk per unit of imports, MacLachlan et al. (2022) found that the number of establishments has remained relatively constant over the past century because of substantial increases in overall import levels and diversification of the origins of imports across regions, which exposed the U.S. to new source species pools.
MacLachlan et al. (2022) suggested that APHIS should target biosecurity resources to the specific commodity-country pairs associated with a higher relative risk of introducing additional insect species.
Recent studies are taking a welcome new stance: looking at links between introductions of non-native plant and insect species. I first raised this approach a year ago. Studies by teams led by Doug Tallany and Sara Lalk [Lalk et al.; articles by Tallamy] agree that:
Non-native plants – some of which are invasive – are altering ecosystems across broad swaths of North America and the impacts are insufficiently understood.
The invasive plant problem will get worse because non-native species continue to be imported, planted … and to invade.
Plant-insect interactions are the foundation of food webs – they transfer energy captured by plants through photosynthesis to other trophic levels, plus play a major role as pollinators. Consequently, changes to a region’s flora will have repercussions throughout ecosystems.
Dr. Tallamy studies the response of herbivorous insects to non-native woody plants – not just invasive plants, but also non-native plants deliberately planted as crops or ornamentals, or in forestry. Introduced plants have completely transformed the composition of plant communities in both natural and human-dominated ecosystems world-wide. The impacts can be significant: Burghardt et al. found that 75% of North American lepidopteran species and 93% of specialist species were found exclusively on native plant species.
monarch butterfly on milkweed; photograph by Jim Hudgins, USFWS
Lalk and colleagues studied the relationships between individual species of invasive woody plants and the full range of arthropod feeding guilds – pollinators, herbivores, twig and stem borers, leaf litter and soil organisms. They decry the absence of data on the complex interactions between invasive woody plants and arthropod communities at a time when invasive shrubs and trees are so widespread and causing considerable ecological damage. (See the blog for their specific research recommendations.)
Nor is the impact of non-native plants on insect fauna limited to North America. Outhwaite et al. found that the combination of climate warming and intensive agriculture is associated with reductions of almost 50% in the abundance and 27% in the number of species within insect assemblages relative to levels in less-disturbed habitats with lower rates of historical climate warming. These patterns were particularly clear in the tropics (perhaps partially because of the longer history of intensive agriculture in temperate zones). They found that high availability of nearby natural habitat (that is, native plants) can mitigate these reductions — but only in low-intensity agricultural systems.
Recognizing that plant diversity drives global patterns of insect invasion, Liebhold et al. (2023) compared various factors associated with numbers of invasive insect species in 44 land areas.They determined that the numbers of established non-native insect species are primarily driven by diversity of plants – both native and non-indigenous. Other factors, e.g., land area, latitude, climate, and insularity, strongly affect plant diversity; thus they influence insect diversity as a secondary impact. When I blogged about this study, I noted that the article appeared more than four years earlier, but has apparently had little influence on either policy formulation governing plant introductions or pest risk analysis applied to insects or pathogens that might be introduced. I suggested that we need a separate analysis of whether fungi, oomycetes, nematodes, and other pathogens show the same association with plant diversity in the receiving environment.
Studies of plant-insect relationships continue to be published. I welcome this!
Bonnamour et al. (2023) builds on the earlier studies. They also found that the presence of non-native plant species was a better predictor of insect invasions than such more widely discussed socioeconomic variables as trade volumes generally or even trade in plant products. However, detection of the associated insect invasions occurs years after detection of the plant invasions. Indeed, numbers of established non-native insect species corresponded more closely to plant introduction volumes in 1900 than current or recent import volumes.
Bonnamour et al. note that while the insect taxa that respond most directly to the non-native plant diversity are those that rely on those plants as hosts, pollinators, and plant visitors, over time those non-native herbaceous insects support introduced predators and parasites also.
Because of the “invasion debt” associated with that lag, Bonnamour et al. estimate that newly detected insect invasions will increase by 35% worldwide as a result of only recent plant introductions. They differentiate this “invasion debt” from “future invasions”, meaning the actual introduction of additional species resulting from future trade activities.
The model developed by Bonnamour et al. points to the highest numbers of newly introduced insect species occurring in areas with less capacity to deal with bioinvasions. Thus, the Afrotropics are anticipated to receive 869 new insect species, or a 10-fold increase over the number currently known to be established in the region. The Neotropics are projected to be invaded by 809 insect species, also a 10-fold increase. The Indomalayan region will probably detect 776 new insect species, a startling 20-fold increase. In reality, the “invasion debt” might not be quite this severe, since – as Bonnamour et al. note several times – the low numbers of introduced insects currently reported for these tropical regions probably partially reflect limited sampling. They note that already a high proportion of insect species intercepted by biosecurity services on imports arriving from Africa and South America are not yet recorded as established in the exporting regions.
Although both the European Palearctic and Australasia have already received many non-native insect species, their “invasion debt” is relatively high: 417 species for Europe, 317 species for Australasia.
The Neotropics are expected to be the greatest source of insect invasions in the future (904 exported species), followed by the European Palearctic (732 species).
Bonnamour et al. did not include non-native plant species used in agriculture, forestry, or ornamental horticulture. As noted above, these widespread deliberate plantings also affect insect fauna and higher trophic layers.
The greatest number of recorded insect introductions so far are in the Nearctic, Oceania (primarily Hawaii), Europe, and Australasia. While this imbalance is probably caused in part by the significantly limited sampling of non-native insect species in the Asian Palearctic and tropics, it is also true that these regions have received the majority of plant introductions through 1900. This factor has changed in the century since then; many non-native plant species have been recorded in the Afrotropics, Oceania, and Asia.
Eucalyptus plantation in Kwa-Zulu-Natal, South Africa; Kwa-Zulu-Natal Dept. of Transportation
Bonnamour et al. offer several potential explanations for the lag in detecting introduced insects compared to detecting introduced plants. First, it might be necessary for non-native host plants to reach a threshold of abundance before the associated insects are able to establish and spread. Second, reaching that threshold might require repeated introductions of the insect’s host plant species. Third, since only some of the imported plants are transporting insects, repeated imports of host plants might be necessary for the insect to achieve sufficient numbers to establish. Fourth, while their analysis included all non-native insect species, only some insect feeding guilds – herbivores and pollinators – are probably directly facilitated by introduced host plants. Fifth, plant species’ presence tends to be more quickly recorded than insects’ presence. Indeed, MacLaughlin et al. reported a median delay of 80 years between establishment and discovery of plant-feeding Hemiptera. This suggests that the actual time lag between plant and insect establishments might be shorter than the period discussed in Bonnamour et al.
Many insects from the European Palearctic have been introduced to the Nearctic; fewer insects have been introduced in the opposite direction. There is no consensus on the explanation. Thirty years ago Mattson et al. argued that there might be fewer niches for non-native insects in Europe due to the lower host plant diversity in this region caused by the Pleistocene/Holocene glaciations. On the other hand, more plant species from the European Palearctic to the Nearctic than the opposite.
Bonnamour et al. call for further research on:
1) time lags at the scale of individual insect species with their host plants.
2) effects of non-native plants used in agriculture, forestry, or ornamental horticulture.
3) whether time lags between plant and insect invasions vary among taxonomic groups, feeding guilds, or among regions.
4) effect of non-native plant abundance, rather than just species richness, on non-native insect establishment.
Recommendations
Writers about interactions of non-native plant species and insect introductions make a common plea: limit the introduction and spread of non-native plants in order to prevent future invasions of both plants and insects. Bonnamour et al. suggest including the risk of insect introductions in plant invasion risk screening tools. Earlier, the Tallamy and Lalk teams called for ending widespread planting of non-native plants.
USDA Secretary Tom Vilsack
Will policy-makers accept this advice?
I believe that these same interaction of plant host and “pest” introductions presumably applies to pathogens, too. I reiterate my frequent complaint that regulators have not responded to two or more decades of criticism of the failures of the international phytosanitary system re: insect and pathogen introductions via the international nursery trade. Examples include Brasier 2008; Liebhold el. al. 2012; Santini et al. 2013; Roy et al. 2014; Eschen et al. 2015; Jung et al. 2015; Meurisse et al. 2019; O’Hanlon et al. 2021.
As I have said earlier, I appreciate that some scientists are trying to reduce scientific uncertainty about the invasive potential of pathogens native to regions other than North America; I refer here to Jiri Hulcr (see Li et al.), Mech, and Schultz. Many more such studies are needed, addressing potential impacts on a wider variety of North American host trees and shrubs.
The late (& very much lamented!) Gary Lovett of the Cary Institute had advocated halting imports of plants that are congenerics of important North American tree species, in order to minimize the risk that pests that damage those genera will be introduced.
In January I suggested that at the global level we need:
National agricultural agencies, stakeholders, FAO & International Plant Protection Convention (IPPC) should consider amending the IPPC requirement that scientists identify a disease’s causal agents before regulating it. Experience shows that this policy virtually guarantees that pathogens will continue to enter, establish, & damage natural and agricultural environments.
National governments & FAO / IPPC should fund greatly expanded research to identify microbes resident in regions that are important sources of origin for traded plants, vulnerability of hosts in importing countries, and new technologies for detecting pathogens (e.g., molecular tools, volatile organic compounds [VOCs]).
Researchers & agencies should expand international “sentinel plants” networks; incorporate data from forestry plantations, urban plantings, etc. of non-native trees.
NPPOs should adopt regulations that apply the “systems approach” or HACCP programs outlined in ISPM#36. I had discussed these approaches in my Fading Forests III report – link at end of this blog.)
I suggested further that Americans need to
Evaluate the efficacy of current regulations – that is, implementing NAPPRA & Q-37 revision. This evaluation should be based on AQIM data, not port interception data. It should include arthropods, fungal pathogens, oomycetes, bacteria, viruses, nematodes. It should include threats to U.S. tropical islands (Hawai`i, Puerto Rico, Guam, etc.) which are centers of plant endemism.
Apply existing programs (e.g., NAPPRA, Clean Stock Network, post-entry quarantine) to strictly regulate trade in plant taxa most likely to transport pests that threaten our native plants; e.g., plants belonging to genera shared between North American trees & plants on other continents.
Recognize that plant nurseries are incubators for microbial growth, hybridization, and evolution; require nurseries to adopt sanitary operation procedures regardless of whether they sell in inter-state or intra-state commerce
SOURCES
Bonnamour, A., R.E. Blake, A.M. Liebhold, H.F. Nahrung, A. Roques, R.M. Turner, T. Yamanaka, and C. Bertelsmeier. 2023. Historical plant intros predict current insect invasions. PNAS 2023 Vol. 120 No. 24 e2221826120 https://doi.org/10.1073/pnas.2221826120
Burghardt, K. T., D. W. Tallamy, C. Philips, and K. J. Shropshire. 2010. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 1(5):art11. doi:10.1890/ES10-00032.
Lalk, S. J. Hartshorn, and D.R. Coyle. 2021. IAS Woody Plants and Their Effects on Arthropods in the US: Challenges and Opportunities. Annals of the Entomological Society of America, 114(2), 2021, 192–205 doi: 10.1093/aesa/saaa054
Li, Y., C. Bateman, J. Skelton, B. Wang, A. Black, Y-T. Huang, A. Gonzalez, M.A. Jusino, Z.J. Nolen, S. Freeman, Z. Mendel, C-Y. Chen, H-F. Li, M. Kolařík, M. Knížek, J-H. Park, W. Sittichaya, T-H.
Pham, S. Itoo, M. Torii, L. Gao, A.J. Johnson, M. Lur, J. Sun, Z. Zhang, D.C. Adams, J. Hulcr. 2022. Pre-invasion assessment of exotic bark beetle-vectored fungi to detect tree-killing pathogens. https://apsjournals.apsnet.org/doi/full/10.1094/PHYTO-01-21-0041-R
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Liebhold, A.M., T. Yamanaka, A. Roques, S. August, S.L. Chown, E.G. Brockerhoff & P. Pyšek. 2018. Plant diversity drives global patterns of insect invasions. Sci Rep8, 12095 (2018). https://doi.org/10.1038/s41598-018-30605-4
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Mattson, W. J., P. Niemela, I. Millers, and Y. Ingauazo. 1994. Immigrant phytophagous insects on woody plants in the United States and Canada: an annotated list. USDA For. Ser. Gen. Tech. Rep. NC-169, 27 pp.
Mech, A.M., K.A. Thomas, T.D. Marisco, D.A. Herms, C.R. Allen, M.P. Ayres, K.J.K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, and P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol. 2019 Nov; 9(21): 12216-12230.,
Richard, M., D.W. Tallamy and A.B. Mitchell. 2019. Intro plants reduce species interactions. Biol Invasions https://doi.org/10.1007/s10530-018-1876-z
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions.
Tallamy, D.W., D.L. Narango and A.B. Mitchell. 2020. Ecological Entomology (2020), DOI: 10.1111/een.12973 Do NIS plants contribute to insect declines? Conservation Biology DOI: 10.1111/j.1523-1739.2009.01202.x
Uden, D.R, A.M. Mech, N.P. Havill, A.N. Schulz, M.P. Ayres, D.A. Herms, A.M. Hoover, K.J. K. Gandhi, R.A. Hufbauer, A.M. Liebhold, T.D. M., K.F. Raffa, K.A. Thomas, P.C. Tobin, C.R. Allen. 2023. Phylogenetic risk assessment is robust for forecasting the impact of European insects on North American conifers. Ecological Applications. 2023; 33:e2761.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
black locust – one of the most widespread invasive tree species on Earth; photo via Flickr
In recent years there has been an encouraging effort to examine bioinvasions writ large see earlier blogs re: costs of invasive species – here and here. One of these products is the Routledge Handbook of Biosecurity and Invasive Species (full citation at end of this blog). I have seen only the chapter on bioinvasion in forest ecosystems written by Sitzia et al. While they describe this situation around the globe, their examples are mostly from Europe.
Similar to other overviews, this article re-states the widely-accepted attribution of rising numbers of species introductions to globalization, especially trade. In so doing, Sitzia et al. assert that the solution is not to curtail trade and movement of people, but to improve scientific knowledge with the goal of strengthening biosecurity and control programs. As readers of this blog know, I have long advocated more aggressive application of stronger restrictions on the most high-risk pathways. Still, I applaud efforts to apply science to risk assessment.
Sitzia et al. attempt to provide a global perspective. They remind readers that all major forest ecosystems of Earth are undergoing significant change as a result of conversion to different land-uses; invasion by a wide range of non-native introduced species—including plants, insects, and mammals; and climate change. These change agents act individually and synergistically. Sitzia et al. give greater emphasis than other writers to managing the tree component of forests. They explain this focus by asserting that forest management could be either the major disturbance favoring spread of non-native species or, conversely, the only way to prevent further invasions. They explore these relationships with the goal of improving conservation of forest habitats.
Japanese stiltgrass invasion; photo by mightyjoepye via Flickr
Sitzia et al. focus first on plant invasions. They contend that – contrary to some expectations – plants can invade even dense forests despite competition for resources. They cite a recent assessment by Rejmánek & Richardson that identified 434 tree species that are invasive around Earth. Many of these species are from Asia, South America, Europe, and Australia. These non-native trees can drive not only changes in composition but also in conservation trajectories in natural forests. However, the example they cite, Japanese stilt grass (Microstegium vimineum) in the United States, is not a tree! Sitzia et al. note that in other cases it is difficult to separate the impacts of management decisions, native competitive species, and non-native species.
Sitzia et al. note that plant invasions might have a wide array of ecological impacts on forests. They attempt to distinguish between
“drivers” of environmental change – including those with such powerful effects that they call them “transformers”;
“passengers” whose invasions are facilitated by other changes in ecosystem properties; and
“backseat drivers” that benefit from changes to ecosystem processes or properties and cause additional changes to native plant communities.
An example of the last is black locust (Robinia pseudoacacia). This North American tree has naturalized on all continents. It is a good example of the management complexities raised by conflicting views of an invasive species’ value, since it is used for timber, firewood, and honey production.
Sitzia et al. then consider invasions by plant pathogens. They say that these invasions are one of the main causes of decline or extirpations in tree populations. I applaud their explicit recognition that even when a host is not driven to extinction, the strong and sudden reduction in tree numbers produces significant changes in the impacted ecosystems.
American chestnut – not extinct but ecological role gone; photo by F.T. Campbell
Sitzia et al. contend that social and economic factors determine the likelihood of a species’ transportation and introduction. Specifically, global trade in plants for planting is widely recognized as being responsible for the majority of introductions. Introductions via this pathway are difficult to regulate because of the economic importance (and political clout) of the ornamental plants industry, large volumes of plants traded, rapid changes in varieties available, and multiple origins of trade. As noted above, the authors seek to resolve these challenges by improving the scientific knowledge guiding biosecurity and control programs. In the case of plant pathogens, they suggest adopting innovative molecular techniques to improve interception efficiency, esp. in the case of latent fungi in asymptomatic plants.
The likelihood that a pathogen transported to a new region will establish is determined by biogeographic and ecological factors. Like other recent studies, Sitzia et al. attempt to identify important factors. They name a large and confusing combination of pathogen- and host-specific traits and ecosystem conditions. These include the fungus’ virulence, host specificity, and modes of action, reproduction, and dispersal, as well as the host’s abundance, demography, and phytosociology. A key attribute is the non-native fungus’ ability to exploit micro-organism-insect interactions in the introduced range. (A separate study by Raffa et al. listed Dutch elm disease as an example of this phenomenon.) I find it interesting that they also say that pathogens that attack both ornamental and forest trees spread faster. They do not discuss why this might be so. I suggest a possible explanation: the ornamental hosts are probably shipped over wide areas by the plant trade.
surviving elms in an urban environment; photo by F.T. Campbell
Sitzia et al. devote considerable attention to bioinvasions that involve symbiotic relationships between bark and ambrosia beetles and their associated fungi. These beetles are highly invasive and present high ecological risk in forest ecosystems. Since ambrosia beetle larvae feed on symbiotic fungi carried on and farmed by the adults inside the host trees, they are often polyphagous. Bark beetles feed on the tree host’s tissues directly, so they tend to develop in a more restricted number of hosts. Both can be transported in almost all kinds of wood products, where they are protected from environmental extremes and detection by inspectors. Sitzia et al. specify the usual suspects: wood packaging and plants for planting, as ideal pathways. These invasions threaten indigenous species by shifting the distribution and abundance of certain plants, altering habitats, and changing food supplies. The resulting damage to native forests induces severe alterations of the landscape and causes economic losses in tree plantations and managed forests. The latter losses are primarily in the high costs of eradication efforts – and their frequent failure.
Eucalyptus plantation in Kwa-Zulu-Natal, South Africa; photo by Kwa-Zulu-Natal Department of Transportation
Perhaps their greatest contribution is their warning about probable damage caused by invasive forest pests in tropical forests. (See an earlier blog about invasive pests in Africa.) Sitzia et al. believe that bark and ambrosia beetles introduced to tropical forests threaten to cause damage of the same magnitude as climate change and clear cutting, but there is little information about such introductions. Tropical forests are exposed to invading beetles in several ways:
1) A long history of plant movement has occurred between tropical regions. Sitzia et al. contend that the same traits sought for commercial production contribute to risk of invasion.
2) Logging and conversion of tropical forests into plantation forestry and agriculture entails movement of potentially invasive plants to new areas. Canopies, understory plant communities, and soils are all disturbed. Seeds, insects, and pathogens can be introduced via contaminated equipment.
3) Less developed nations are often at a disadvantage in managing potential invasion. Resources may be fewer, competing priorities more compelling, or potential threats less obvious.
Sitzia et al. call for development of invasive species management strategies that are relevant to and realistic for less developed countries. These strategies must account for interactions between non-native species and other aspects of global environmental change. Professional foresters have a role here. One clear need is to set out practices for dealing with conflicts between actors driven by contrasting forestry and conservation interests. These approaches should incorporate the goals of shielding protected areas, habitat types and species from bioinvasion risk. Sitzia et al. also discuss how to address the fact that many widely used forestry trees are invasive. (See my earlier blog about pines planted in New Zealand.)
planted forest in Sardinia, Italy; photo by Torvlag via Flickr
In Europe, bark beetle invasions have damaged an estimated ~124 M m2 between 1958 and 2001. Sitzia et al. report that the introduction rate of non-native scolytins has increased sharply. As in the US, many are from Asia. They expect this trend to increase in the future, following rising global trade and climate change. Southern – Mediterranean – Europe is especially vulnerable. The region has great habitat diversity; a large number of potential host trees; and the climate is dry and warm with mild winters. The region has a legacy of widespread planting of non-native trees which are now important components of the region’s economy, history and culture. These include a significant number of tree species that are controversial because they are – or appear to be – invasive. Thus, new problems related to invasive plants are likely to emerge.
Noting that different species and invasion stages require different action, Sitzia et al. point to forest planning as an important tool. Again the discussion centers on Europe. Individual states set forest policies. Two complications are the facts that nearly half of European forests are privately owned; and stakeholders differ in their understanding of the concept of “sustainability”. Does it mean ‘sustainable yield’ of timber? Or providing multiple goods and services? Or sustaining evolution of forest ecosystems with restrictions on the use of non-native species? Resolving these issues requires engagement of all the stakeholders.
Sitzia et al. say there has recently been progress. The Council of Europe issued a voluntary Code of Conduct on Invasive Alien Trees in 2017 that provides guidelines on key pathways. A workshop in 2019 elaborated global guidelines for the sustainable use of non-native tree species, based on the Bern Convention Code of Conduct on Invasive Alien Trees. The workshop issued eight recommendations:
Use native trees, or non-invasive non-native trees;
Comply with international, national, and regional regulations concerning non-native trees;
Be aware of the risk of bioinvasion and consider global change trends;
Design and adopt tailored practices for plantation site selection and silvicultural management;
Promote and implement early detection and rapid response programs;
Design and adopt practices for invasive non-native tree control, habitat restoration, and for dealing with highly modified ecosystems;
Engage with stakeholders on the risks posed by invasive NIS trees, the impacts caused, and the options for management; and
Develop and support global networks, collaborative research, and information sharing on native and non-native trees.
SOURCE
Sitzia, T., T. Campagnaro, G. Brundu, M. Faccoli, A. Santini and B.L. Webber. 2021 Forest Ecosystems. in Barker, K. and R.A. Francis. Routledge Handbook of Biosecurity and Invasive Species. ISBN 9780367763213
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
This blog asks YOU!!! to support funding for some of the key USDA programs. This blog focuses on USDA’s Animal and Plant Health Inspection Service (APHIS). APHIS is responsible for preventing introduction of pests that harm agriculture, including forests; and for immediate efforts to eradicate or contain those pests that do enter. While most port inspections are carried out by the Department of Homeland Security Bureau of Customs and Border Protection, APHIS sets the policy guidance. APHIS also inspects imports of living plants.
Please help by contacting your members of the House and Senate Appropriations Committees. I provide a list of members – by state – at the end of this blog. APHIS is funded by the House and Senate Appropriations Subcommittees on Agriculture and Related Agencies. These Subcommittees have scheduled hearings on the topic and I’ve drafted written testimony for them. I expect CISP will be joined by additional members of the Sustainable Urban Forest Coalition in signing the testimony. You can add the crucial voice of constituent’s support.
I will blog soon about funding for USDA’s Forest Service (USFS) – I don’t yet have necessary information to suggest specific funding levels.
Your letter or email need be no more than a couple paragraphs. To make the case for greater funding, feel free to pick-and-choose from the information that follows. Your greatest impact comes from speaking specifically about what you know and where you live.
These are the specific dollar amounts we’d like you to ask for. The rationale for each is below.
Appropriations for APHIS programs (in $ millions)
Program
FY 2022 (millions)
FY 2023
FY 2024 Pres.’ request
Our ask
Tree & Wood Pest
$61
$63
$64
$65 M
Specialty Crops
$210
$216
$222
$222 M
Pest Detection
$28
$29
$30
$30 M
Methods Development
$21
$23
$23
$25 M
The Costs of Introduced Pests
Introduced pests threaten many forest products and services benefitting all Americans, including wood products, wildlife habitat, carbon sequestration, clean water and air, storm water management, lower energy costs, improved health, aesthetic enjoyment, and related jobs. Already, the 15 most damaging non-native pests threaten at least 41% of forest biomass in the “lower 48” states. In total, these 15 species have caused an additional annual conversion of live biomass to dead wood at a rate similar in magnitude to that attributed to fire (5.53 TgC per year for pests versus 5.4 to 14.2 TgC per year for fire) [Fei et al.; full citation at end of blog; see also earlier].
tanoaks killed by SOD; Oregon Department of Forestry photo
These pests also impose significant costs that are borne principally by municipal governments and homeowners. As more pests have been accidentally introduced over time, these costs have risen. A study published last year [Hudgins et al.] projected that by 2050 1.4 million street trees in urban areas and communities will be killed by introduced insect pests. Municipalities on the forefront include Milwaukee and Madison Wisconsin; the Chicago area; Cleveland; and Baltimore, Towson, and Salisbury, Maryland. Removing and replacing these trees is projected to cost cities $30 million per year. Additional urban trees – in parks, on homeowners’ properties, and in urban woodlands – are also expected to die and require removal and replacement.
Pathways of Introduction
Tree-killing pests are linked to the international supply chain. Many pests—especially the highly damaging wood-borers like emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot hole borers, and redbay ambrosia beetle—arrive in inadequately treated crates, pallets, and other forms of packaging made of wood. Other pests—especially plant diseases like sudden oak death and sap sucking insects like hemlock woolly adelgid—come on imported plants. Some pests take shelter, or lay their eggs, in or on virtually any exposed hard surface, such as steel, decorative stone, or shipping containers.
infested wood from a crate; Oregon Department of Agriculture photo
Wood Packaging
Imports from Asia have historically transported the most damaging pests, e.g., Asian longhorned beetle, emerald ash borer, redbay ambrosia beetle, and the invasive shot hole borers. For decades goods from Asia have dominated imports. As of February 2022, U.S. imports from Asia were running at a rate of 20 million shipping containers per year. A recent analysis [Haack et al.; see also here] indicates that at least 33,000 of these shipping containers, perhaps twice that number, are carrying a tree-killing pest. These facts have led scientists to project [Leung et al.] that by 2050, the number of non-native wood-boring insects established in the US could triple. Hudgins et al. say the greatest damage would occur if an Asian wood-boring insect that attacks maples or oaks were introduced. Such a pest could kill 6.1 million trees and cost American cities $4.9 billion over 30 years. The risk would be highest if this pest were introduced to the South – and U.S. southern ports are receiving more direct shipments from Asia after the expansion of the Panama Canal in 2016. https://www.nivemnic.us/?m=202207
After introduction of the ALB, APHIS acted to curtail further introductions in wood packaging from China. First – in 1998 – APHIS required China to treat its wood packaging. Second, it worked with foreign governments to develop the International Standard for Phytosanitary Measures (ISPM) #15. The U.S. and Canada began phasing in ISPM#15 in 2005 with full implementation in 2006. Under ISPM#15, all countries shipping goods to North America must treat their wood packaging according to specified protocols with the goal of “significantly reducing” the risk that pests will be present.
However, as I have often blogged [see blogs under “wood packaging” category on this site] ISPM#15 has fallen short. Haack et al. found that as recently as 2020, 0.22% [1/5th of 1 percent] of the shipping containers entering the U.S. were infested by a tree-killing insect. This equates to tens of thousands of containers harboring tree-killing insects.
Worse, the data indicate that our trade partners’ compliance with the rules has deteriorated; the “approach rate” of pest-infested wood packaging fell in 2005-2006, but has since gone back up.
The most troubling offender is China. Although required since 1998 to treat its wood packaging, China consistently has one of the highest pest approach rates: it was 0.73% [or ¾ of 1%] during the 2010-2020 period. This is three times the global average for the period. Since China supplied 40.7% of U.S. imports in 2022 [Szakonyi], or 5,655,000 containers. Thus China alone might be sending to the U.S. 30,000 containers infested with tree-killing insects. These pests threaten our urban, rural, and wildland forests and reduce forest productivity, carbon sequestration, the rural job base, water supplies and quality, and many other ecosystem services.
ISPM#15 falls short at the global level. The fact that a pallet or crate bears the mark indicating that it complies with ISPM#15 has not proved to be reliable.
You might ask your Member of Congress or Senators to ask APHIS what steps it will take to correct the problem of Chinese non-compliance. (Remind him or her that that the Asian longhorned beetle, emerald ash borer, and many other insects of so-far lesser impact were introduced in wood packaging from China.
Asian longhorned beetle
Remind them also that the Department of Homeland Security’s Bureau of Customs and Border Protection has twice enhanced its enforcement of wood packaging rules. In 2017 it began penalizing importers of non-compliant wood packaging under Title 19 United States Code (USC) §1595a(b) or under 19 USC §1592. In 2021, it incorporated the wood packaging requirements into its voluntary C-TPAC program.)
You might also urge them to ask APHIS what steps it is taking at the global level to improve the efficacy of ISPM#15 – or to replace it if necessary to ensure that pests are not being introduced.
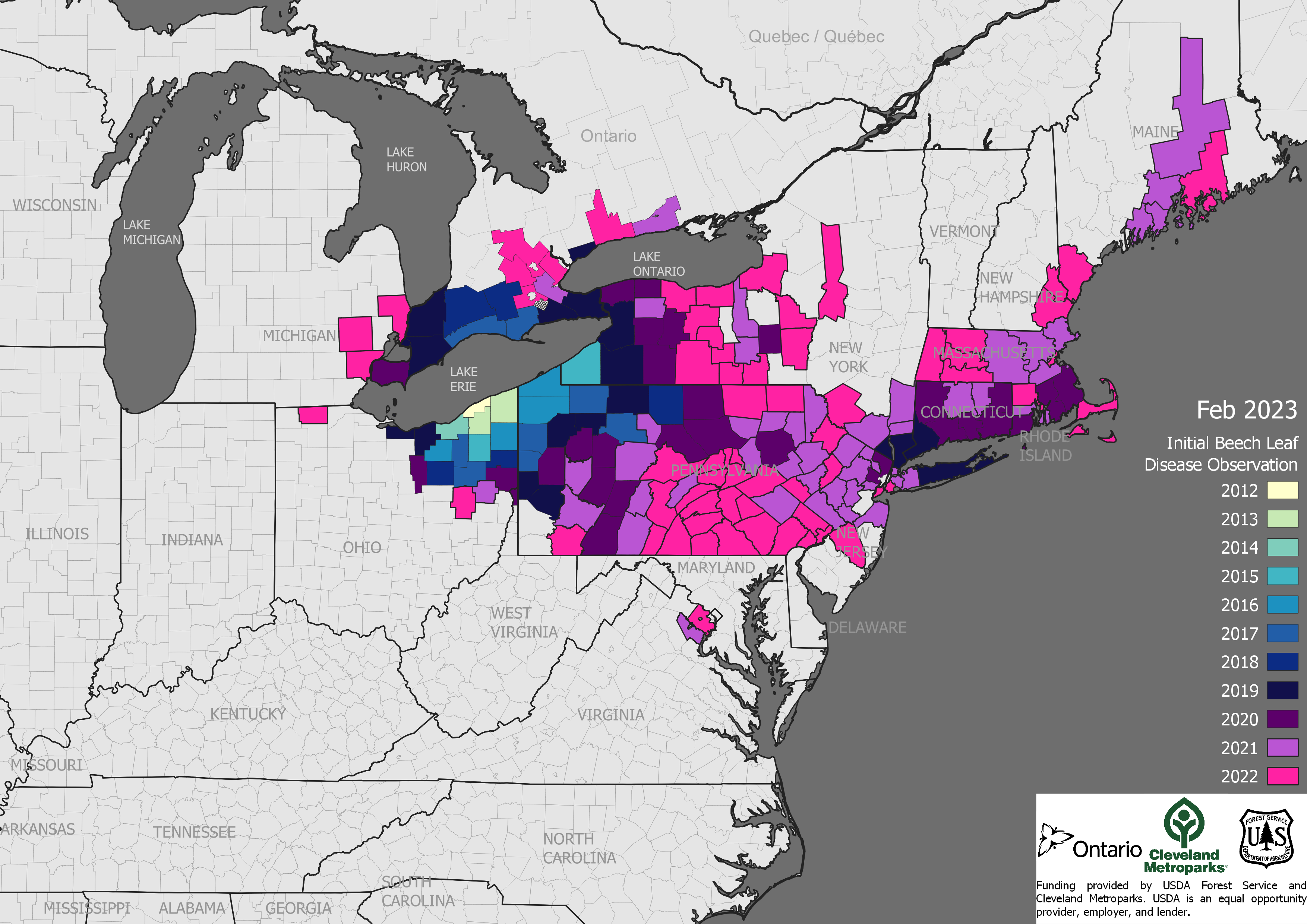
spread of beech leaf disease
Imported Plants (“Plants for Planting”)
Some pest types—especially plant diseases like sudden oak death and sap-sucking insects like hemlock woolly adelgid—come on imported plants. The U.S. imported about 5 billion plants in 2021 [MacLachlan]. Recent introductions probably via this pathway include several pathogens — Phytophthoras, rapid ʻōhiʻa death in Hawai`i, beech leaf disease (established from Ohio to Maine), and boxwood blight. Insects have also been introduced on imported plants recently; one example is the elm zigzag sawfly (present in North Carolina, Virginia, and New York and Ontario). https://www.nivemnic.us/?p=4115
An analysis of data from 2009 [Liebhold et al.] found that approximately 12% of plant shipments were infested by a pest. This pest approach rate is more than 50 times higher than the 0.22% approach rate for wood packaging. APHIS has adopted several changes to its phytosanitary system for imported plants in the decade since 2009. A few studies have been published, but they have focussed on insects and excluded pathogens. We have noted that pathogens continue to be introduced via the plant trade. Therefore, please ask your Member or Senators to ask APHIS to facilitate an independent analysis of the efficacy of the agency’s current phytosanitary programs to prevent introductions of pests on important plants, with an emphasis on introductions of plant pathogens.
APHIS is responsible for preventing spread of the SOD pathogen, Phytophthora ramorum, through trade in nursery plants. In recent years California has had few detections in nurseries and little expansion in forests – but the situation suggests that this good news is probably more the result of the drought than of program efficacy. In cooler, wetter conditions in Oregon and Washington, detections in nurseries and alarming detections in the forest or plantings continue.
In 2022, the APHIS SOD Program supported detection and regulatory activities in 25 states. P. ramorum was detected at 18 establishment, 12 of which were first-time detections. The California nursery regulatory program – which is funded by APHIS – saw reduced funding in 2022. We think these cuts are unwise since this year’s very wet winter will probably lead to a new disease outbreaks. Programs in Oregon and Washington continue to detect infestations in additional retailers brought in by plants bought from other nurseries. Washington responded to four separate “trace forward” incidents, one involving more than 160 residential sites. Clearly, the federal-state program is not succeeding in eradicating P. ramorum from nurseries. Please suggest that your Congressperson and Senators ask APHIS what steps it is taking to improve the efficacy of the SOD program.
SOD-infected rhodoendron on plants in Indiana; photo by Indiana Department of Natural Resources
In the East, P. ramorum was found in three of 65 streams sampled in 10 states in 2022 (reaching across the Southeast from Mississippi through North Carolina, plus Texas, Maryland, Pennsylvania, and Illinois). One stream is troubling: a first-time detection in South Carolina, with no obvious nursery source. Since stream sampling began, P. ramorum has been detected from eight streams in four states, Alabama, Mississippi, North Carolina, and now South Carolina. The pathogen has been present in some of these streams for more than 10 years.
Oregon faces particularly high risks. Three of the four known strains of P. ramorum are established in Oregon forests. One of them, the EU1 lineage, is more aggressive than the NA1 clonal lineage already present in forests. In addition, the EU1 strain might facilitate sexual reproduction of the pathogen, thus exacerbating Oregon’s struggle to contain the disease.
As we know, introduced pests do not stay in the cities where they first arrived — they spread! Often that spread is facilitated by our movement of firewood, plants, or outdoor household goods such as patio furniture.
The beech trees so important to wildlife conservation in the Northeast are under attack by two pathogens and at risk to an insect. Most alarming is the spread – in a dozen years! — of beech leaf disease DMF from Ohio to Maine. A leaf-feeding weevil is spreading south in eastern Canada. Please suggest that your Member or Senators to ask APHIS what steps it is taking to prevent the weevil’s introduction to the U.S.
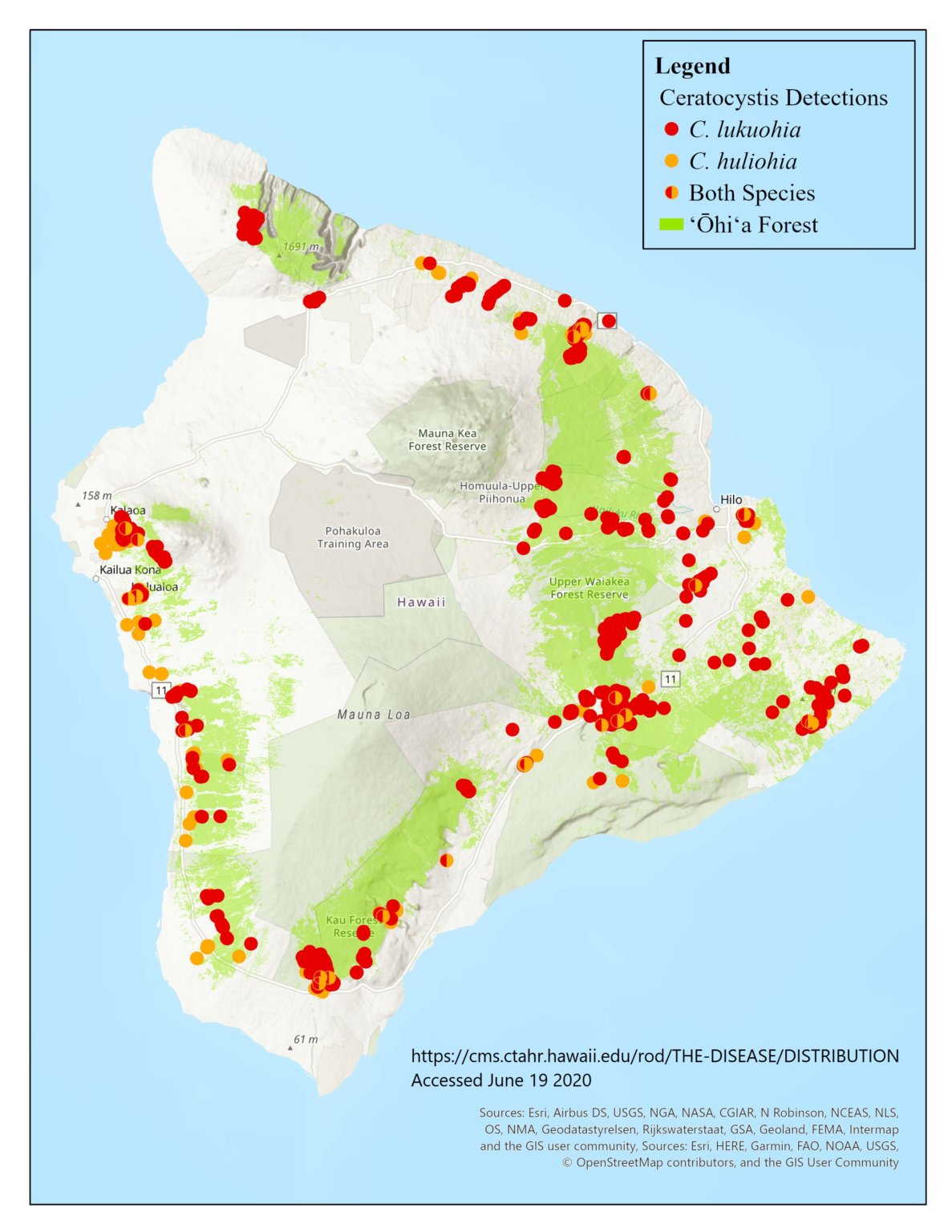
‘Ōhi‘a trees make up 80% of the biomass of forests in both wet and dry areas of the Hawaiian archipelago. It is under attack by two diseases caused by introduced pathogens first detected in 2010. ‘Ōhi‘a forests support more threatened and endangered species than any other forest system in the U.S. They also play a uniquely important role in providing other ecosystem services, including water supplies.
Asking for the Money Pest Problems Deserve
To respond effectively to these pests and to the others that will be introduced in coming years, the key APHIS programs identified above must have adequate funds. The funding levels I request – and hope you will support – are lower than I would wish, but everyone expects the Congress to refuse significant increases in funding (see table at beginning of this blog).
The Tree and Wood Pests account supports eradication and control efforts targeting principally the ALB and spongy (= gypsy) moth. Eradicating the ALB normally receives about two-thirds of the funds. The programs in Massachusetts, New York, Ohio, and South Carolina must continue until eradication succeeds.
Oregon detected the EAB in 2022. Although the state and Portland have been preparing for a decade for this eventuality, there will still be significant impacts. Four percent of Portland’s street trees are ash – more than 9,000 trees. Young ash constitute three percent of young trees in parks. Loss of Oregon’s ash will also have severe ecosystem impacts. In Willamette Valley wetlands, ash constitutes up to 100% of the forest trees. Washington and California are also concerned. Indeed, the Hudgins study identified Seattle and Takoma as likely to lose thousands of ash trees. The numerous ash in riparian forests, windbreaks, and towns of North Dakota are also at risk since the EAB is established in South Dakota, Minnesota, and Manitoba.
APHIS manages damaging pests introduced on imported plants or other items through its Specialty Crops program. The principal example is its efforts to prevent spread of the SOD pathogen through the interstate trade in nursery plants. We noted above that this program is not as successful as it should be. We support the Administration’s request for $222 million; however, you might suggest that your Member or Senator urge APHIS to allot adequate funding under this budget line to management of SOD, rapid ʻōhiʻa death pathogens in Hawai`i, and beech leaf disease and elm zig-zag sawfly in the East.
The Pest Detection program is key to the prompt detection of newly introduced pests that is critical to successful pest eradication or containment. The “Methods Development” program enables APHIS to improve development of essential detection and eradication tools.
The Administration’s request include a $1 million emergency fund. This is far below the level needed to respond when a new pest is discovered. Funding constraints have hampered APHIS’ response to past pest incursions.
Please note that many of the members of the Agriculture Appropriations Subcommittee are from states where non-native pests are probably not top of mind. It is important that everyone that knows about these threats communicate with your Member/Senators!!
Members of House or Senate Subcommittees that Fund APHIS
(Names of Senators are italicized)
STATE
MEMBER
APHIS APPROP
HOUSE
SENATE
AK
Lisa Murkowski
X
AL
Jerry Carl Katie Britt
X
X
X
Calif
Barbara Lee David Valadao Josh Harder Diane Feinstein
X X X
X X X
X
FL
Debbie Wasserman Scultz Scott Franklin
X X
X X
GA
Sanford Bishop
X
X
ID
Mike Simpson
X
IL
Lauren Underwood
X
X
KS
Jerry Moran
X
X
KY
Mitch McConnell
X
X
LA
Julia Letlow Ashley Hinson
X X
X X
MD
Andy Harris Chris Van Hollen
X
X
X
ME
Chellie Pingree Susan Collins
X X
X
X
MI
John Moolenaar Gary Peters
X X
x
X
MN
Betty McCollum
X
X
MS
Cindy Hyde-Smith
X
X
MT
Jon Tester Ryan Zinke
X
X
X
NB
Deb Fischer
X
ND
John Hoeven
X
X
NM
Martin Heinrich
X
X
NV
Mark Amodei
X
OH
Marcy Kaptur
X
X
OR
Jeff Merkley
X
X
X
PA
Guy Reschenthaler
X
X
RI
Jack Reed
X
TX
Michael Cloud Jake Ellzey
X
X X
UT
Chris Stewart
X
VA
Ben Cline
X
X
WA
Dan Newhouse Derek Kilmer
X
X X
WV
Shelly Moore CapitoJoe Manchin
X
X X
WI
Mark Pocan Tammy Baldwin
X X
X
X
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice .2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Hudgins, E.J., F.H. Koch, M.J. Ambrose, and B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ 2014; doi:10.1890/130311
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. Frontiers in Ecology.
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Szakonyi, M. 2023. Sourcing shift from China pulls US import share to more than a decade low.
spotted lanternfly; photo by Stephen Ausmus, USDA; establishment facilitated by extent of invasion by its preferred host, Ailanthus
Seven coauthors (full citation at end of blog) compared various factors associated with numbers of invasive insect species in 44 land areas.These ranged from small oceanic islands to entire continents in different world regions, Liebhold et al. determined that the numbers of established non-native insect species are primarily driven by diversity of plants, including both native and non-indigenous. Other factors, e.g., land area, latitude, climate, and insularity, strongly affect plant diversity. Through this mechanism these factors affect insect diversity as a secondary impact.
Seven coauthors (full citation at end of blog) compared various factors associated with numbers of invasive insect species in 44 land areas.These ranged from small oceanic islands to entire continents in different world regions, Liebhold et al. determined that the numbers of established non-native insect species are primarily driven by diversity of plants, including both native and non-indigenous. Other factors, e.g., land area, latitude, climate, and insularity, strongly affect plant diversity. Through this mechanism these factors affect insect diversity as a secondary impact.
At large spatial scales [greater than 10 km2], regions supporting more diverse plant communities offer greater opportunities for herbivore colonization. Thus, plant diversity promotes invasion through the “facilitation effect”. Since most insects – including most of those introduced to naïve ecosystems – are herbivores, a greater number of possible foods is a clear advantage. Those insects that prey on herbivores benefit by plant diversity indirectly.
Non-native coral tree, Erythrina, in Hawai`i; photo by Forrest and Kim Starr; did wide planting of exotic Erythrina facilitate invasion by Erythrina gall wasp?
At smaller spatial scales, plant diversity might impair the ability of insects to locate hosts because of the “dilution effect”. I have been asking for decades why so few of the Eurasian insects established in eastern North America have not also established along the Pacific coast from Oregon into British Columbia. The region has a plant-friendly climate and almost every plant species from temperate climates is grown there in cultivation. Perhaps the non-native plants – while numerous enough to become invaders themselves – are still sufficiently scarce or dispersed to impair introduced insects’ locating an familiar host?
According to the Smithsonian Institution, Hawai`i has approximately 2,499 taxa of flowering plants and 222 taxa of ferns and related groups. The native flora of the United States includes about 17,000 species of vascular plants; at least 3,800 non-native species of vascular plants are recorded as established outside cultivation. I don’t know how many non-native plant species are in cultivation.
horticultural viburnum invading riparian forest in Fairfax County, VA. photo by F.T. Campbell; did the widespread presence of many non-native viburnum species facilitate establishment of the viburnum leaf beetle?
I note that this article appeared more than four years ago. However, its important findings do not appear to have been integrated into either policy formulation governing plant introductions or pest risk analysis applied to insects or pathogens that might be introduced. (Indeed, we probably need a separate analysis of whether fungi, oomycetes, nematodes, and other pathogens show the same association with plant diversity in the receiving environment.)
How do we – government agencies, academics, conservation organizations, plant industry representatives — use this information to help curtail introductions of plant pests? Can it be integrated into APHIS’ NAPPRA process?
SOURCE
Liebhold, A.M., T. Yamanaka, A. Roques, S. August, S.L. Chown, E.G. Brockerhoff & P. Pyšek. 2018. Plant diversity drives global patterns of insect invasion. www.nature.com/scientificreports/
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
carnation (Dianthus caryophyllus) photo by Noordzee23 via Wikimedia; carnations have been big sellers for 250 years
Plants sold in nurseries directly influence urban landscapes by providing gardens and other habitats that support humans and birds, insects, and other organisms. Doug Tallamy, though, has described ways that non-native plants fall short in providing habitat for native wildlife. Of course, non-native plants also indirectly influence natural landscapes by acting as a major source of invasive species. [see blog – includes links to regional invasive plant lists; and here] Imported plants also can carry non-native insects and pathogens – about which I blog frequently! To review these blogs, scroll down below the archives to the “categories” section and click on “plants as pest vectors”.
Now Kinlock, Adams, and van Kleunen (full citation at the end of this blog) have published a new paper that sheds more light on these issues. They analyzed the ornamental plants sold in US nurseries over 225 years (from 1719 to 1946). Their database, drawn from an earlier publication by Adams (see Sources at end of blog), included records of 5,098 ornamental vascular plant species offered for sale by 319 US nursery catalogs published over this period.
They note that present-day urban yards in the continental United States are planted in a diverse array of plants and the plants are predominately non-native species. Also, there is relatively little variation in species planted from one region to another, especially when compared to regional variation in natural areas). These patterns reflect the history of US horticulture.
Seventy percent (3,587) of the 5,098 ornamental vascular plant species offered by the 319 nurseries over those 200 years were non-native to the continental United States. They believe that the number of non-native species offered for sale has probably continued to increase in the 70 years since their study ended. They cite a study showing that 91% of tree species sold by nurseries in southern California during the 20th and early 21st centuries were not native to that state. A similar figure comes from a study of cultivated plants grown in Minneapolis–Saint Paul. There 66% of plants were non-native. (Kinlock, Adams, and van Kleunen note that 70% of species cultivated in yards of five British cities are non-native. In contrast, only 23% of cultivated plants in 18 Chinese cities were non-native.)
Kinlock, Adams, and van Kleunen note that two examples of non-native plants that have become invasive were among most common species available from nurseries beginning in the mid-19th Century: Japanese honeysuckle (Lonicera japonica) was available in 78 nurseries, and Japanese barberry (Berberis thunbergia) in 46 nurseries.
Japanese honeysuckle; photo by Chuck Bargeron, Bugwood
Historical Trends
The earliest commercial horticulture in colonies that became the United States was in the mid-17th Century. It involved imports of Eurasian fruit trees to establish orchards to provide familiar foods. Ornamental horticulture became popular earlier than I expected. Prince Nurseries was established in 1732 in Flushing, NY. It was followed by additional nurseries in New York, Philadelphia, and Massachusetts. Originally these businesses imported Old World nursery stock and seeds – again to provide familiar foods and take advantage of relationships with European contacts.
Nurseries proliferated in the 1820s in the population centers of the Atlantic coast. As people of European ancestry moved west, so did nurseries. Kinlock, Adams, and van Kleunen point out an interesting aspect of these changes: proliferation of both was aided by technology: steamboats, canals, highways, and improved mail service. Before 1800, nearly all nurseries were in the Mid-Atlantic, New England, and South. Nurseries appeared in the Great Lakes region by the 1830s. Expansion of rail lines connected nurseries from coast to coast by the 1870s. By 1890, there were more than 4,500 nurseries across the continent.
California, Florida, and Oregon are now the states with the most horticultural operations and sales (as of 2019).
The types of plants offered for sale proliferated throughout the 19th Century. The species richness of US nursery flora peaked in the early 20th Century. It decreased in the 1925 – 1946 period, possibly attributable to some combination of war-related interruptions to trade and a shift in gardeners’ focus away from ornamentals to vegetables. Another factor was adoption of international and interstate phytosanitary regulations in the early 20th Century. The post-World War II economic boom led to a new diversification of US nursery flora. In one study, a Los Angeles nursery experienced the largest increase in species richness during 1990–2011. They believe this increase was probably matched across the country. Global plant collection and importation mediated by US botanical gardens and nurseries remain active.
planting of Eucalyptus seedlings in California during 1980s; National Archives photo
Over time, nursery floras in the various regions became more similar to each other. The floras of Mid-Atlantic and New England nurseries differed before 1775, then became similar. Nurseries in the Great Lakes region also shifted toward offerings in neighboring regions. Later, nurseries in the South and West also began offering a higher proportion of species commonly sold across the continent. The nursery floras of Great Lakes and Great Plains regions were consistently similar. Still, the flora in Western nurseries still retain some unique aspects. California is the only state with a Mediterranean climate. Nurseries there sought adapted plant species, especially from an entirely new source — Australasia. (The authors note that Acacia and Eucalyptus genera, while important in California horticulture, are invaders in Mediterranean zones worldwide.) One might expect the need for plants in the Southwest to be drought-tolerant would prompt a unique nursery flora. However, the ubiquity of irrigation since the late 19th Century has blunted this necessity. Still, nursery flora in the desert biome had the most phylogenetic uniformity. The article does not discuss pressure to choose xeriscapes or otherwise adjust to current water shortages.
Pinus mogu – sought for xerescapes; photo by Krysztof Ziamk Kenraiz via Wikimedia
Growing Importance of Non-Native Species – Especially from Asia
Kinlock, Adams, and van Kleunen define “native” species as those native to the state in which it is sold; “adventive” species as native to the continental United States but not the specific state; and non-native or alien species as not native to the continental United States.
Applying these definitions, the proportion of native species in nursery flora has been consistently around 30-40% — except during the American Revolution. It rose to 70% in catalogs or advertisements published from 1775 to 1799. The authors do not speculate whether this reflected jingoism or interruptions in trade. The proportion of plant species that were adventive was 4% in the earliest period, then rose to 13% with improved transportation.
A large proportion of the native species offered in the late 18th and early 19th centuries were grown for export to Europe (think John Bartram).
Rhodendron maximum; sent to Europe by John Bartram (& invasive in Great Britain and Ireland!); photo from Pl@ntNet.identify
Throughout the 19th and 20th centuries, plants from new regions of the world with unique genetic lineages became increasingly available. Until the mid-19th Century, most non-native plants came from Europe and Eurasia. Beginning in 1850, plants native to temperate Asia composed an increasing percentage of non-native nursery flora. In the period 1900 – 1924, 19% of the ornamental nursery flora originated from temperate Asia. By the next period, 1925 – 1946, this percentage rose to 20.8%.At the same time, North American species (including some from Mexico, Canada, or Alaska) composed 21.9% of the nursery flora. (see graph).
% of species from various origins; North America – medium blue; temperate Asia – dark pink; Europe – tan; Eurasia – fuscia; Southern America – blue-green; Africa – yellow-green; Americas – olive
Plants from East Asia were particularly desirable for both biological and social reasons. First, because of climatic similarities between the two regions, East Asian plants thrived in the eastern United States. Second, popular ornamental genera had higher species richness in East Asia. Important social or cultural factors were a growing fascination with Japanese and Chinese-style gardens: forced “opening” of access to those countries in the 1840s and 1850s; and plant collecting expeditions sponsored by British and American institutions and private collectors. In 1898, the US Department of Agriculture established the Section of Seed & Plant Introduction; its purpose was to collect and cultivate economically useful non-native plants from throughout South America and Asia.
As I noted above, diversity of species in nursery offerings reached a peak in the first years of the 20th Century, concurrent with the first wave of US-sponsored plant collections; indeed, 70 species that were first listed after 1911 in their dataset were introduced by the USDA introduction program between 1912 -1942.
Commodore Perry in Japan; Library of Congress
Counter-pressures and Counter-measures
There were counter pressures during this period that – as mentioned above—probably contributed to a decline in plant introductions in later years. In the 1890s, several US states began requiring inspection of imported plant materials (spurred by plant disease outbreaks caused by spread of San Jose scale from California).
Congress adopted the Plant Quarantine Act in 1912; USDA implemented it through stringent regulations issued in 1919 (Quarantine-37). I have already noted interruption of trade associated with WWI and WWII. Kinlock et al. don’t mention the Great Depression that intervened, but I think it played a role, too. On the other hand, Q-37 was relaxed to target particular species or regions based on pest risk analysis. The article says the relaxation began in the 1930s, but I believe it actually was during the 1970s; see Liebhold et al. 2012. I have blogged several times about how well the current regulations – including the “NAPPRA” program – prevent introductions of invasive plants or damaging plant pests. To review these blogs, scroll down below the archives to the “categories” section and click on “plants as pest vectors”.
dogwood anthracnose; photo by Robert Anderson, USFS; via Bugwood
SOURCES
Adams, D.W. 2004. Restoring American Gardens: An encyclopedia of heirloom ornamental plants. Timber Press
Kinlock, N.L., D.W. Adams, M. van Kleunen. 2022. An ecological and evolutionary perspective of the historical US nursery flora. Plants People Planet. 2022;1–14. wileyonlinelibrary.com/journal/ppp3
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
APHIS is seeking stakeholder input to its new strategic plan to guide the agency’s work over the next 5 years.
The strategic plan framework is a summary of the draft plan; it provides highlights including the mission and vision statements, core values, strategic goals and objectives, and trends or signals of change we expect to influence the agency’s work in the future. APHIS is seeking input on the following questions:
Are your interests represented in the plan?
Are there opportunities for APHIS to partner with others to achieve the goals and objectives?
Are there other trends for which the agency should be preparing?
Are there additional items APHIS should consider for the plan?
range of American beech – should APHIS be doing more to protect it from 3 non-native pests?
Comments must be received by July 1, 2022, 11:59pm (EST).
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
monarch butterfly on swamp milkweed; photo by Jim Hudgins, USFWS
I have been impressed recently by two groups of scientists who are trying to broaden understanding of the impacts of invasive plants by examining the interactions of those plants with insects. As they note, herbivorous insects are key players in terrestrial food webs; they transfer energy captured by plants through photosynthesis to other trophic levels. This importance has been recognized since Elton first established the basic premises of food webs (1927) [Burghardt et al.; full citation at end of blog] Arthropods comprise significant members of nearly every trophic level and are especially important as pollinators. If introduced plants cause changes to herbivore communities, there will probably be effects on predators, parasites, and other wildlife through multitrophic interactions [Lalk et al.; Tallamy, Narango and Mitchell].
[I briefly summarize the findings of a third group of scientists at the end of this blog. The third group looks at the interaction between agriculture – that is, planting of non-native plants! – and climate change.]
One approach to studying this issue, taken by Douglas Tallamy of the University of Delaware and colleagues, is to look at the response of herbivorous insects to NIS woody plants fairly generally. They integrate their studies with growing concern about the global decline in insect populations and diversity. They note that scientists have focused on light pollution, development, industrial agriculture, and pesticides as causes of these declines. They decry the lack of attention to disruption of specialized evolutionary relationships between insect herbivores and their native host plants due to widespread domination by non-indigenous plants [Richard, Tallamy and Mitchell].
In their studies, Tallamy and colleagues consider not just invasive plants, but also non-native plants deliberately planted as crops or ornamentals, or in forestry. They point out that such introduced plants have completely transformed the composition of plant communities in both natural and human-dominated ecosystems around the globe. At least 25% of the world’s planted forests are composed of tree species not native to their locale. At least one-sixth of the globe is highly vulnerable to plant invasions, including biodiversity hotspots [Richard, Tallamy and Mitchell].
A different approach, taken by Lalk and colleagues, is more closely linked to concern about impacts of the plants themselves. They have chosen to pursue knowledge about relationships between individual species of invasive woody plants and the full range of arthropod feeding guilds – pollinators, herbivores, twig and stem borers, leaf litter and soil organisms. In so doing, they decry the general absence of data.
Both teams agree that:
Invasive plants are altering ecosystems across broad swaths of North America and the impacts are insufficiently understood.
The invasive plant problem will get worse because non-native species continue to be imported and planted. (Reminder: the Tallamy team considers impacts of deliberate planting as well as bioinvasion.)
Plant-insect interactions are the foundation of food webs, so changes to them will have repercussions throughout ecosystems.
Tallamy team
Non-native plants have replaced native plant communities to a greater or lesser extent in every North American biome – including anthropogenic landscapes [Burghardt]. The first trophic level in suburban and urban ecosystems throughout the U.S. is dominated by plant species that evolved in Southeast Asia, Europe, and South America [Tallamy and Shropshire]. Abundant non-native plants not only dominate plant biomass; they also reduce native plant taxonomic, functional and phylogenetic diversity and heterogeneity. Thus, they indirectly alter the abundance of native insects [Burghardt; Richard, Tallamy and Mitchell].
I think these articles might actually underestimate the extent of these impacts. While Richard, Tallamy and Mitchell report that more than 3,300 species of non-native plants are established in continental U.S., years ago Rod Randall said that more than 9,700 non-native plant species were naturalized in the U.S. (probably includes Hawai` i. The Tallamy team cites USDA Forest Service data showing 9% of forests in the southeast are invaded by just 33 common invasive plant species [Richard, Tallamy and Mitchell], I have cited that and other sources showing even greater extents of plant invasion in the east and here; other regions and here.
The Tallamy team has conducted several field experiments that demonstrate that the presence of non-native plants suppress numbers and diversity of native lepidopteran caterpillars. These non-native woody plants have not replaced the ecological functions of the native plants that used to support insect populations. This is true whether or not the non-native plants are deliberately planted or are invading various ecosystems on their own. [Richard, Tallamy and Mitchell]. (Of course, they expect plant invasions to grow; they note that some of the many ornamental species that are not yet invasive will become so.)
The result is disruption of the ecological services delivered by native plant communities, including the focus of their studies: plants’ most fundamental contribution to ecosystem function: generation of food for other organisms [Burghardt].
They note that plants’ relationship to insects differs depending on the insects’ feeding guilds — folivores, wood eaters, detritivores, pollinators, frugivores, and seed-eaters; and among herbivores with different mouthparts — chewing or sucking; and as host plant specialists or generalists. They decry studies that fail to recognize these differences [Tallamy, Narango, and Mitchell].
The Tallamy team explores why insect populations decline among non-native plants. That is,
1) Do insects directly requiring plant resources have lower fitness when using non-native plants; do they not recognize them as viable host plants; or do they avoid them altogether?
2) Are reductions in numbers of specialist herbivores mitigated by generalists? A decade of research shows that both specialists and generalists decline.
The team’s studies focus on lepidopteran larvae (caterpillars). Insect herbivores are both the largest taxon of primary consumers and extremely important in transferring energy captured by plants through photosynthesis to other trophic levels [Burghardt]. In addition, insects with chewing mouthparts are typically more susceptible to defensive secondary metabolites contained in leaves than are insects with sucking mouthparts that tap into poorly defended xylem or phloem fluids [Tallamy, Narango and Mitchell].
A study by Burghardt et al. found that 75% of all lepidopteran species and 93% of specialist species were found exclusively on native plant species. Non-native plants that were in the same genus as a native plant often supports a lepidopteran community that is a similar but depauperate subset of the community found on its native congener. In fact, the insect abundance and species richness supported by non-native congeners of native species was reduced by 68%.
A meta-analysis of 76 studies by other scientists found that, with few exceptions, caterpillars had higher survival and were larger when reared on native host plants. Plant communities invaded by non-native species had significantly fewer Lepidoptera and less species richness. In three of eight cases examined, non-native plants functioned as ecological traps, inducing females to lay eggs on plants that did not support successful larval development. Richard, Tallamy and Mitchell cite as an example the target of many conservation efforts, monarch butterflies (Danaus plexxipus), which fail to reproduce when they use nonnative swallowworts (Vincetoxicum species.) instead of related milkweeds (Asclepias species.).
Tallamy and Shropshire ranked 1,385 plant genera that occur in the mid-Atlantic region by their ability to support lepidopteran species richness. They found that introduced ornamentals are not the ecological equivalents of native ornamentals. This means that solar energy harnessed by introduced plants is largely unavailable to native specialist insect herbivores.
Tallamy, Narango, and Mitchell describe the following patterns:
1) Insects with chewing mouthparts are typically more susceptible to defensive secondary metabolites contained in leaves than are insects with sucking mouthparts that tap into poorly defended xylem or phloem fluids. As a result, sucking insects find novel non-indigenous plants to be acceptable hosts more often. However, there are more than 4.5 times as many chewing (mandibulate) insect herbivores than sucking (haustellate) species. It follows that the largest guild of insect herbivores is also the most vulnerable to non-native plants as well as being the most valuable to insectivores.
native azalea Rhododendron periclymenoides; photo by F.T. Campbell
2) Woody native species, on average, support more species of phytophagous insects than herbaceous species.
3) Although insects are more likely to accept non-native congeners or con-familial species as novel hosts, non-native congeners still reduced insect abundance and species richness by 68%.
4) Host plant specialists are less likely to develop on evolutionarily novel non-indigenous plants than are insects with a broader diet. There are far more specialist species than generalists, so generalists will not prevent serious declines in species richness and abundance when native plants are replaced by non-indigenous plants. In addition, non-native plants cause significant reductions in species richness and abundance even of generalists. In fact, generalists are often locally specialized on particular plant lineages and thus may function more like specialists than expected.
5) Any reduction in the abundance and diversity of insect herbivores will probably cause a concomitant reduction in the insect predators and parasitoids of those herbivores – although few studies have attempted to measure this impact beyond spiders, which are abundant generalists. The vast majority of parasitoids are highly specialized on particular host lineages.
6) Studies comparing native to non-native plants must avoid using native species that support very few phytophagous insects as their baseline, e.g., in the mid-Atlantic region tulip poplar trees (Liriodendron tulipifera) and Yellowwood (Cladrastus kentuckea).
7) Insects that feed on well-defended living tissues such as leaves, buds, and seeds are less likely to be able to include non-native plants in their diets than are insects that develop on undefended tissues like wood, fruits, and nectar. Although this hypothesis has never been formally tested, they note the ease with which introduced wood borers – emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot-hole borers, redbay ambrosia beetle, Sirex woodwasp (all described in profiles posted here — have become established in the US.
palamedes swallowtail Papilio palamedes; photo by Vincent P. Lucas; this butterfly depends on redbay, a tree decimated by laurel wilt disease vectored by the redbay ambrosia beetle
Lalk and Colleagues
As noted, Lalk and colleagues have a different frame; they focus on individual introduced plant species rather than starting from insects. They also limit their study to invasive plants. The authors say there is considerable knowledge about interactions between invasive herbaceous plants and arthropod communities, but less re: complex interactions between invasive woody plants and arthropod communities, including mutualists (e.g., pollinators), herbivores, twig- and stem-borers, leaf-litter and soil-dwelling arthropods, and other arthropod groups.
They ask why this knowledge gap persists when invasive shrubs and trees are so widespread and causing considerable ecological damage. They suggest the answer is that woody invaders rarely encroach on high-value agricultural systems and some are perceived as contributing ecosystem services, including supporting some pollinators and wildlife.
Lalk and colleagues seek to jump-start additional research by summarizing what is currently known about invasive woody plants’ interactions with insects. They found sufficient data about 11 species – although even these data are minimal. They note that all have been cultivated and sold in the U.S. for more than 100 years. All but one (mimosa) are listed as a noxious weed by at least one state; two states (Rhode Island and Georgia) don’t have a noxious weed list. None of the 11 is listed under the federal noxious weed statute.
Ailanthus altissima
Illustrations of how minimal the existing information is:
Tree-of-heaven (Ailanthus altissima) is noted to be supporting expanded populations of the Ailanthus webworm moth (Atteva aurea), which is native to Central America; and to be the principal reproductive host for SLF (Lycorma delicatua) https://www.dontmovefirewood.org/pest_pathogen/spotted-lanternfly-html/
Chinese tallow (Triadica sebifera) is thought to benefit both native generalist bee species and non-indigenous European honeybees (Apis mellifera).
Chinese privet (Ligustrum sinense) appears to suppress populations of butterflies, bees, and beetles.
Lalk and colleagues then review what is known about interactions between individual invasive plant species in various feeding guilds. They point out that existing data on these relationships are scarce and sometimes contradictory.
They believe this is because interactions vary depending on phylogenetic relationships, trophic guild, and behavior (e.g., specialized v. generalist pollinator). Arthropods can be “passengers” of a plant invasion. That is, they can be affected by that invasion, with follow-on effects to other arthropods in the community. Also, arthropods can be “drivers” of invasion, increasing the success of the invasive plants.
They then summarize the available information on various interactions. For example, they note that introduced plants can compete with native plants in attracting pollinators, causing cascading effects. Or they can increase pollination services to native plants by attracting additional pollinators.
They note that herbivore pressure on invasive plants can have important impacts on growth, spread, and placement within food webs. They note that these cases support the “enemy release hypothesis”, although they think there are probably additional driving mechanisms.
Lalk and colleagues note that most native twig- and stem-borers (Coleoptera: Buprestidae, Curculionidae, Cerambycidae; Hymenoptera: Siricidae) are not considered primary pests but that some of our most damaging insect species are wood borers (see above).
Some of these borers are decomposers; in that role, they are critical in nutrient cycling.
Arthropods in leaf litter and soil also serve important roles in the decomposition and cycling of nutrients, which affects soil biota, pH, soil nutrients, and soil moisture. They act as a trophic base in many ecosystems. Lalk and colleagues suggest these arthropod communities probably change with plant species due to differences in leaf phytochemistry. They cite one study that found litter community composition differed significantly between litter beneath tree-of-heaven, honeysuckle (Lonicera maackii), and buckthorn (Rhamnus cathartica) compared to litter underneath surrounding native trees.
Recommendations
Both the Tallamy and Lalk teams call for ending widespread planting of non-native plants. Lalk and colleagues discuss briefly the roles of
The nursery industry (including retailers); they produce what sells.
Scientists and educators have not sufficiently informed home and land owners about which species are invasive or about native alternatives.
Private citizens buy and plant what their neighbors have, what they consider aesthetically pleasing, or what is being promoted.
States have not prohibited sale of most invasive woody plants. Regulatory actions are not a straightforward matter; they require considerable time, supporting information, and compromise.
Tallamy team calls for restoration ecologists in the eastern U.S. to consider the number of Lepidopterans hosted by a plant species when deciding what to plant. For example, oaks (Quercus), willows (Salix), native cherries (Prunus)and birches (Betula) host orders of magnitude more lepidopteran species in the mid-Atlantic region than tulip poplar.(Those lepidopteran in turn support breeding birds and other insectivorous organisms.) [Tallamy & Shropshire]
Lalk and colleagues focused on identifying several key knowledge gaps:
How invasive woody plants affect biodiversity and ecosystem functioning
How they themselves function in different habitats.
Do non-native plants drive shifts in insect community composition, and if so, what is that shift, and how does it affect other trophic levels?
How do IAS woody plants affect pollinators?
The authors do not minimize the difficulty of separating such possible plant impacts from other factors, including climate change and urbanization.
Outhwaite et al. (full citation at end of this blog) note that past studies have shown that insect biodiversity changes are driven primarily by land-use change (which is another way of saying planting of non-native species – as Dr. Tallamy and colleagues describe it) and increasingly by climate change. They south to examine whether these drivers interact. They found that the combination of climate warming and intensive agriculture is associated with reductions of almost 50% in the abundance and 27% in the number of species within insect assemblages relative to levels in less-disturbed habitats with lower rates of historical climate warming. These patterns were particularly clear in the tropics (perhaps partially because of the longer history of intensive agriculture in temperate zones). They found that high availability of nearby natural habitat (that is, native plants) can mitigate these reductions — but only in low-intensity agricultural systems.
Outhwaite et al. reiterate the importance of insect species in ecosystem functioning, citing pollination, pest control, soil quality regulation & decomposition. To prevent loss of these important ecosystem services, they call for strong efforts to mitigate climate change and implementation of land-management strategies that increase the availability of natural habitats.
SOURCES
Burghardt, K. T., D. W. Tallamy, C. Philips, and K. J. Shropshire. 2010. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 1(5):art11. doi:10.1890/ES10-00032.
Lalk, S. J. Hartshorn, and D.R. Coyle. 2021. IAS Woody Plants and Their Effects on Arthropods in the US: Challenges and Opportunities. Annals of the Entomological Society of America, 114(2), 2021, 192–205 doi: 10.1093/aesa/saaa054
Richard, M. D.W. Tallamy and A.B. Mitchell. 2019. Introduced plants reduce species interactions. Biol Invasions
Tallamy, D.W., D.L. Narango and A.B. Mitchell. 2020. Ecological Entomology (2020), DOI: 10.1111/een.12973 Do Non-native plants contribute to insect declines?
Tallamy, D.W. and K.J. Shropshire. 2009. Ranking Lepidopteran Use of Native Versus Introduced Plants Conservation Biology, Volume 23, No. 4, 941–947 2009 Society for Conservation Biology DOI: 10.1111/j.1523-1739.2009.01202.x
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm