Help Fight for Money to Protect Forests
This blog asks YOU!!! to support funding for some of the key USDA programs. This blog focuses on USDA’s Animal and Plant Health Inspection Service (APHIS). APHIS is responsible for preventing introduction of pests that harm agriculture, including forests; and for immediate efforts to eradicate or contain those pests that do enter. While most port inspections are carried out by the Department of Homeland Security Bureau of Customs and Border Protection, APHIS sets the policy guidance. APHIS also inspects imports of living plants.
Please help by contacting your members of the House and Senate Appropriations Committees. I provide a list of members – by state – at the end of this blog. APHIS is funded by the House and Senate Appropriations Subcommittees on Agriculture and Related Agencies. These Subcommittees have scheduled hearings on the topic and I’ve drafted written testimony for them. I expect CISP will be joined by additional members of the Sustainable Urban Forest Coalition in signing the testimony. You can add the crucial voice of constituent’s support.
I will blog soon about funding for USDA’s Forest Service (USFS) – I don’t yet have necessary information to suggest specific funding levels.
Your letter or email need be no more than a couple paragraphs. To make the case for greater funding, feel free to pick-and-choose from the information that follows. Your greatest impact comes from speaking specifically about what you know and where you live.
These are the specific dollar amounts we’d like you to ask for. The rationale for each is below.
Appropriations for APHIS programs (in $ millions)
| Program | FY 2022 (millions) | FY 2023 | FY 2024 Pres.’ request | Our ask |
| Tree & Wood Pest | $61 | $63 | $64 | $65 M |
| Specialty Crops | $210 | $216 | $222 | $222 M |
| Pest Detection | $28 | $29 | $30 | $30 M |
| Methods Development | $21 | $23 | $23 | $25 M |
The Costs of Introduced Pests
Introduced pests threaten many forest products and services benefitting all Americans, including wood products, wildlife habitat, carbon sequestration, clean water and air, storm water management, lower energy costs, improved health, aesthetic enjoyment, and related jobs. Already, the 15 most damaging non-native pests threaten at least 41% of forest biomass in the “lower 48” states. In total, these 15 species have caused an additional annual conversion of live biomass to dead wood at a rate similar in magnitude to that attributed to fire (5.53 TgC per year for pests versus 5.4 to 14.2 TgC per year for fire) [Fei et al.; full citation at end of blog; see also earlier].

These pests also impose significant costs that are borne principally by municipal governments and homeowners. As more pests have been accidentally introduced over time, these costs have risen. A study published last year [Hudgins et al.] projected that by 2050 1.4 million street trees in urban areas and communities will be killed by introduced insect pests. Municipalities on the forefront include Milwaukee and Madison Wisconsin; the Chicago area; Cleveland; and Baltimore, Towson, and Salisbury, Maryland. Removing and replacing these trees is projected to cost cities $30 million per year. Additional urban trees – in parks, on homeowners’ properties, and in urban woodlands – are also expected to die and require removal and replacement.
Pathways of Introduction
Tree-killing pests are linked to the international supply chain. Many pests—especially the highly damaging wood-borers like emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot hole borers, and redbay ambrosia beetle—arrive in inadequately treated crates, pallets, and other forms of packaging made of wood. Other pests—especially plant diseases like sudden oak death and sap sucking insects like hemlock woolly adelgid—come on imported plants. Some pests take shelter, or lay their eggs, in or on virtually any exposed hard surface, such as steel, decorative stone, or shipping containers.

Wood Packaging
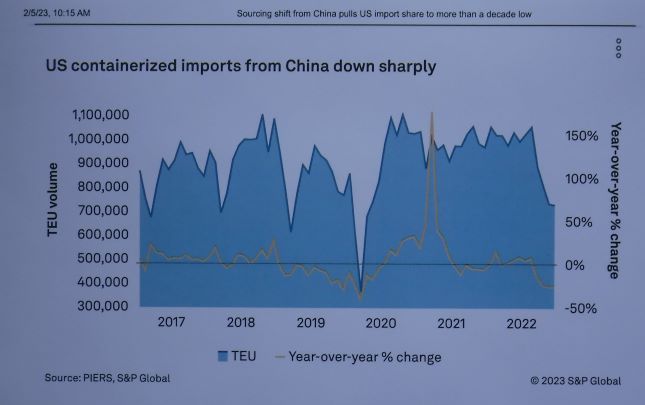
Imports from Asia have historically transported the most damaging pests, e.g., Asian longhorned beetle, emerald ash borer, redbay ambrosia beetle, and the invasive shot hole borers. For decades goods from Asia have dominated imports. As of February 2022, U.S. imports from Asia were running at a rate of 20 million shipping containers per year. A recent analysis [Haack et al.; see also here] indicates that at least 33,000 of these shipping containers, perhaps twice that number, are carrying a tree-killing pest. These facts have led scientists to project [Leung et al.] that by 2050, the number of non-native wood-boring insects established in the US could triple. Hudgins et al. say the greatest damage would occur if an Asian wood-boring insect that attacks maples or oaks were introduced. Such a pest could kill 6.1 million trees and cost American cities $4.9 billion over 30 years. The risk would be highest if this pest were introduced to the South – and U.S. southern ports are receiving more direct shipments from Asia after the expansion of the Panama Canal in 2016. https://www.nivemnic.us/?m=202207
After introduction of the ALB, APHIS acted to curtail further introductions in wood packaging from China. First – in 1998 – APHIS required China to treat its wood packaging. Second, it worked with foreign governments to develop the International Standard for Phytosanitary Measures (ISPM) #15. The U.S. and Canada began phasing in ISPM#15 in 2005 with full implementation in 2006. Under ISPM#15, all countries shipping goods to North America must treat their wood packaging according to specified protocols with the goal of “significantly reducing” the risk that pests will be present.
However, as I have often blogged [see blogs under “wood packaging” category on this site] ISPM#15 has fallen short. Haack et al. found that as recently as 2020, 0.22% [1/5th of 1 percent] of the shipping containers entering the U.S. were infested by a tree-killing insect. This equates to tens of thousands of containers harboring tree-killing insects.
Worse, the data indicate that our trade partners’ compliance with the rules has deteriorated; the “approach rate” of pest-infested wood packaging fell in 2005-2006, but has since gone back up.
The most troubling offender is China. Although required since 1998 to treat its wood packaging, China consistently has one of the highest pest approach rates: it was 0.73% [or ¾ of 1%] during the 2010- 2020 period. This is three times the global average for the period. Since China supplied 40.7% of U.S. imports in 2022 [Szakonyi], or 5,655,000 containers. Thus China alone might be sending to the U.S. 30,000 containers infested with tree-killing insects. These pests threaten our urban, rural, and wildland forests and reduce forest productivity, carbon sequestration, the rural job base, water supplies and quality, and many other ecosystem services.
ISPM#15 falls short at the global level. The fact that a pallet or crate bears the mark indicating that it complies with ISPM#15 has not proved to be reliable.
You might ask your Member of Congress or Senators to ask APHIS what steps it will take to correct the problem of Chinese non-compliance. (Remind him or her that that the Asian longhorned beetle, emerald ash borer, and many other insects of so-far lesser impact were introduced in wood packaging from China.

Remind them also that the Department of Homeland Security’s Bureau of Customs and Border Protection has twice enhanced its enforcement of wood packaging rules. In 2017 it began penalizing importers of non-compliant wood packaging under Title 19 United States Code (USC) §1595a(b) or under 19 USC §1592. In 2021, it incorporated the wood packaging requirements into its voluntary C-TPAC program.)
You might also urge them to ask APHIS what steps it is taking at the global level to improve the efficacy of ISPM#15 – or to replace it if necessary to ensure that pests are not being introduced.
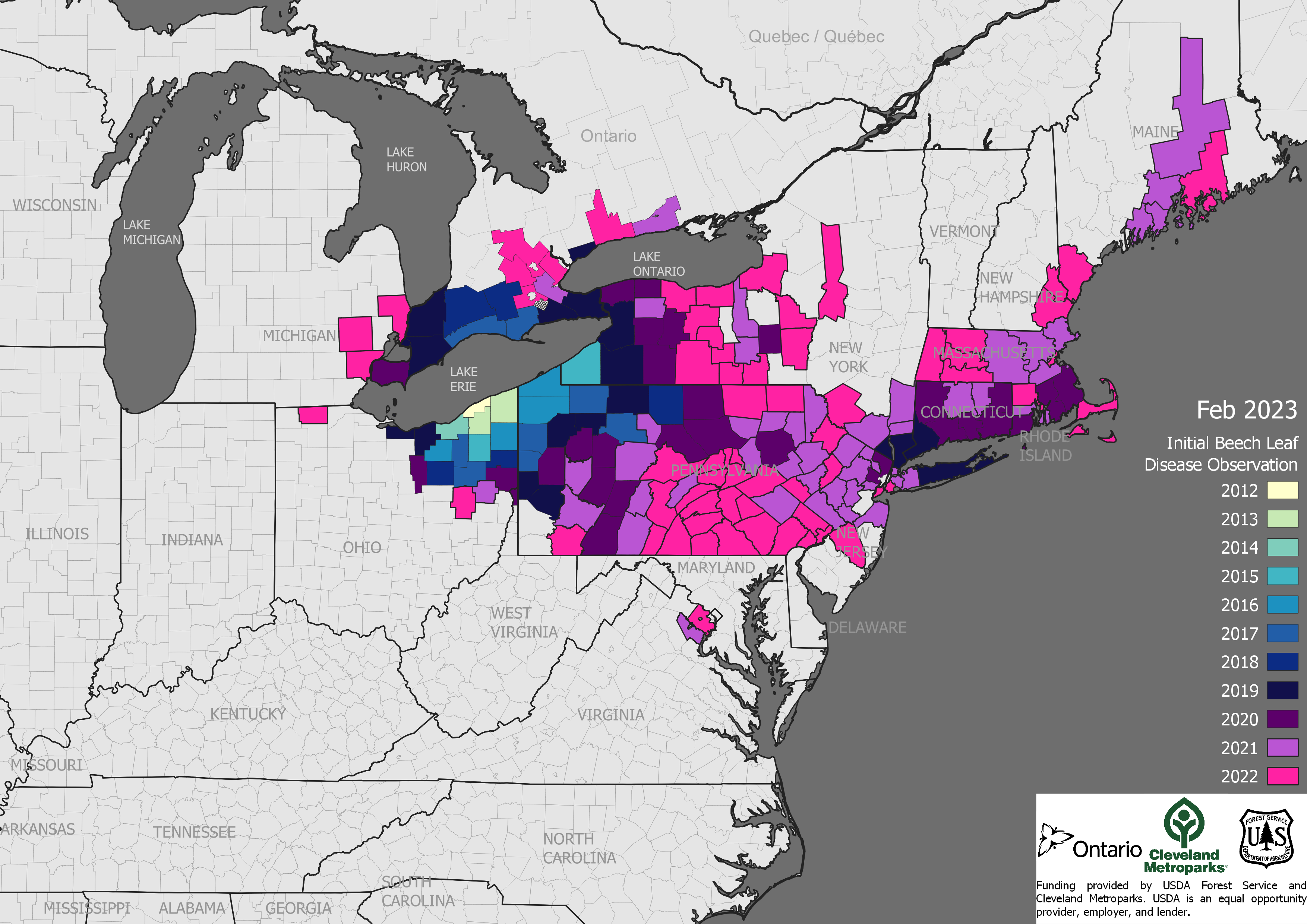
Imported Plants (“Plants for Planting”)
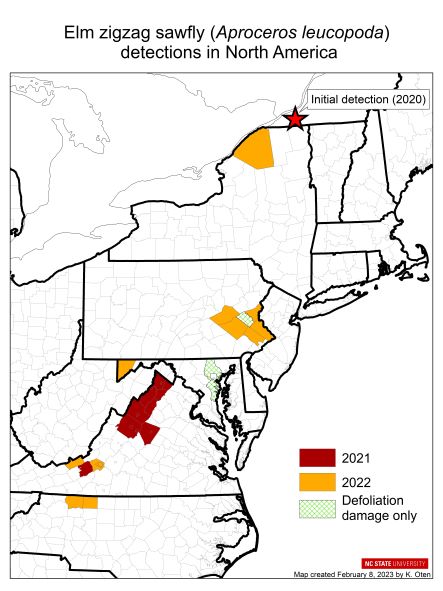
Some pest types—especially plant diseases like sudden oak death and sap-sucking insects like hemlock woolly adelgid—come on imported plants. The U.S. imported about 5 billion plants in 2021 [MacLachlan]. Recent introductions probably via this pathway include several pathogens — Phytophthoras, rapid ʻōhiʻa death in Hawai`i, beech leaf disease (established from Ohio to Maine), and boxwood blight. Insects have also been introduced on imported plants recently; one example is the elm zigzag sawfly (present in North Carolina, Virginia, and New York and Ontario). https://www.nivemnic.us/?p=4115
An analysis of data from 2009 [Liebhold et al.] found that approximately 12% of plant shipments were infested by a pest. This pest approach rate is more than 50 times higher than the 0.22% approach rate for wood packaging. APHIS has adopted several changes to its phytosanitary system for imported plants in the decade since 2009. A few studies have been published, but they have focussed on insects and excluded pathogens. We have noted that pathogens continue to be introduced via the plant trade. Therefore, please ask your Member or Senators to ask APHIS to facilitate an independent analysis of the efficacy of the agency’s current phytosanitary programs to prevent introductions of pests on important plants, with an emphasis on introductions of plant pathogens.
APHIS is responsible for preventing spread of the SOD pathogen, Phytophthora ramorum, through trade in nursery plants. In recent years California has had few detections in nurseries and little expansion in forests – but the situation suggests that this good news is probably more the result of the drought than of program efficacy. In cooler, wetter conditions in Oregon and Washington, detections in nurseries and alarming detections in the forest or plantings continue.
In 2022, the APHIS SOD Program supported detection and regulatory activities in 25 states. P. ramorum was detected at 18 establishment, 12 of which were first-time detections. The California nursery regulatory program – which is funded by APHIS – saw reduced funding in 2022. We think these cuts are unwise since this year’s very wet winter will probably lead to a new disease outbreaks. Programs in Oregon and Washington continue to detect infestations in additional retailers brought in by plants bought from other nurseries. Washington responded to four separate “trace forward” incidents, one involving more than 160 residential sites. Clearly, the federal-state program is not succeeding in eradicating P. ramorum from nurseries. Please suggest that your Congressperson and Senators ask APHIS what steps it is taking to improve the efficacy of the SOD program.

In the East, P. ramorum was found in three of 65 streams sampled in 10 states in 2022 (reaching across the Southeast from Mississippi through North Carolina, plus Texas, Maryland, Pennsylvania, and Illinois). One stream is troubling: a first-time detection in South Carolina, with no obvious nursery source. Since stream sampling began, P. ramorum has been detected from eight streams in four states, Alabama, Mississippi, North Carolina, and now South Carolina. The pathogen has been present in some of these streams for more than 10 years.
Oregon faces particularly high risks. Three of the four known strains of P. ramorum are established in Oregon forests. One of them, the EU1 lineage, is more aggressive than the NA1 clonal lineage already present in forests. In addition, the EU1 strain might facilitate sexual reproduction of the pathogen, thus exacerbating Oregon’s struggle to contain the disease.
As we know, introduced pests do not stay in the cities where they first arrived — they spread! Often that spread is facilitated by our movement of firewood, plants, or outdoor household goods such as patio furniture.
The beech trees so important to wildlife conservation in the Northeast are under attack by two pathogens and at risk to an insect. Most alarming is the spread – in a dozen years! — of beech leaf disease DMF from Ohio to Maine. A leaf-feeding weevil is spreading south in eastern Canada. Please suggest that your Member or Senators to ask APHIS what steps it is taking to prevent the weevil’s introduction to the U.S.
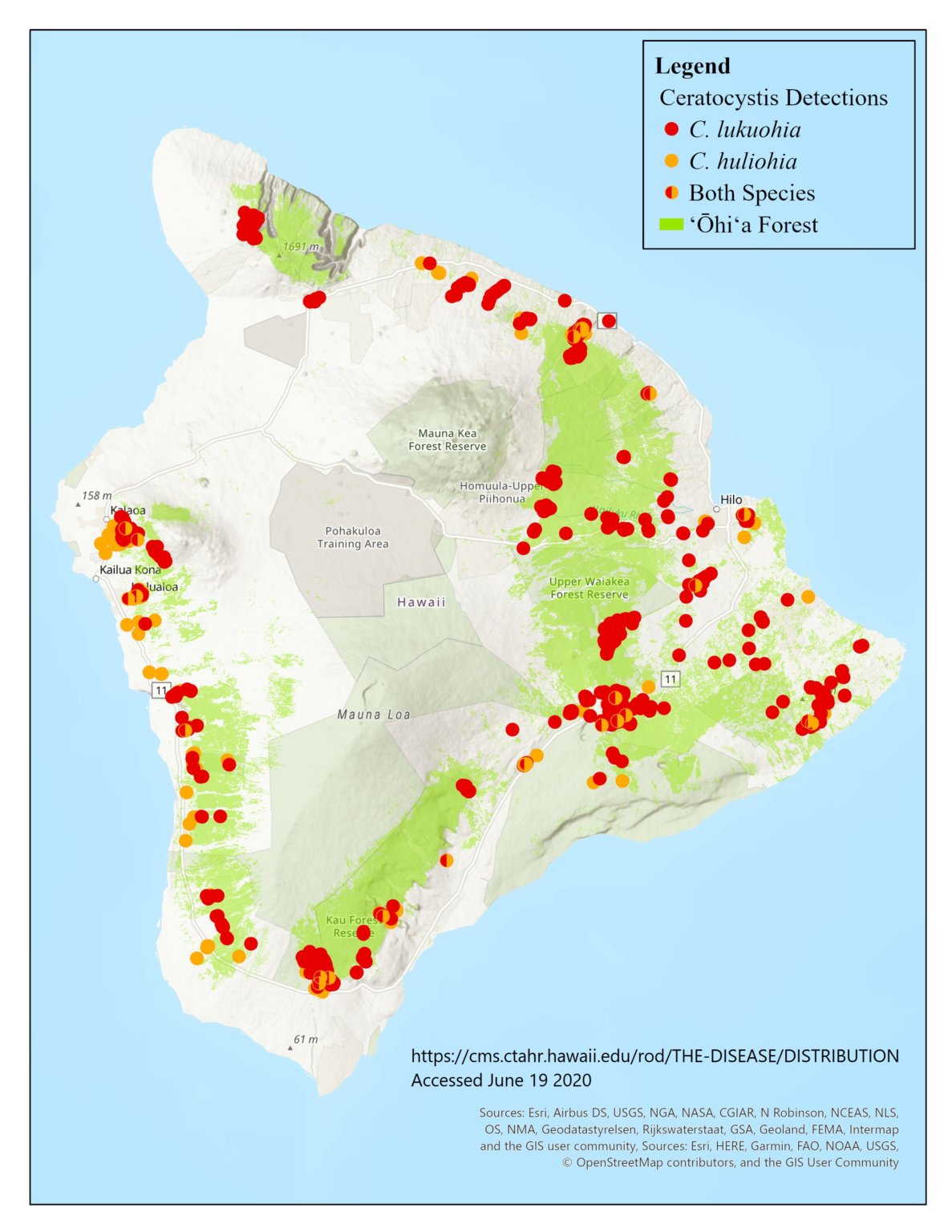
‘Ōhi‘a trees make up 80% of the biomass of forests in both wet and dry areas of the Hawaiian archipelago. It is under attack by two diseases caused by introduced pathogens first detected in 2010. ‘Ōhi‘a forests support more threatened and endangered species than any other forest system in the U.S. They also play a uniquely important role in providing other ecosystem services, including water supplies.
Asking for the Money Pest Problems Deserve
The Tree and Wood Pests account supports eradication and control efforts targeting principally the ALB and spongy (= gypsy) moth. Eradicating the ALB normally receives about two-thirds of the funds. The programs in Massachusetts, New York, Ohio, and South Carolina must continue until eradication succeeds.
Oregon detected the EAB in 2022. Although the state and Portland have been preparing for a decade for this eventuality, there will still be significant impacts. Four percent of Portland’s street trees are ash – more than 9,000 trees. Young ash constitute three percent of young trees in parks. Loss of Oregon’s ash will also have severe ecosystem impacts. In Willamette Valley wetlands, ash constitutes up to 100% of the forest trees. Washington and California are also concerned. Indeed, the Hudgins study identified Seattle and Takoma as likely to lose thousands of ash trees. The numerous ash in riparian forests, windbreaks, and towns of North Dakota are also at risk since the EAB is established in South Dakota, Minnesota, and Manitoba.
APHIS manages damaging pests introduced on imported plants or other items through its Specialty Crops program. The principal example is its efforts to prevent spread of the SOD pathogen through the interstate trade in nursery plants. We noted above that this program is not as successful as it should be. We support the Administration’s request for $222 million; however, you might suggest that your Member or Senator urge APHIS to allot adequate funding under this budget line to management of SOD, rapid ʻōhiʻa death pathogens in Hawai`i, and beech leaf disease and elm zig-zag sawfly in the East.
The Pest Detection program is key to the prompt detection of newly introduced pests that is critical to successful pest eradication or containment. The “Methods Development” program enables APHIS to improve development of essential detection and eradication tools.
The Administration’s request include a $1 million emergency fund. This is far below the level needed to respond when a new pest is discovered. Funding constraints have hampered APHIS’ response to past pest incursions.
Please note that many of the members of the Agriculture Appropriations Subcommittee are from states where non-native pests are probably not top of mind. It is important that everyone that knows about these threats communicate with your Member/Senators!!
Members of House or Senate Subcommittees that Fund APHIS
(Names of Senators are italicized)
| STATE | MEMBER | APHIS APPROP | HOUSE | SENATE |
| AK | Lisa Murkowski | X | ||
| AL | Jerry Carl Katie Britt | X | X | X |
| Calif | Barbara Lee David Valadao Josh Harder Diane Feinstein | X X X | X X X | X |
| FL | Debbie Wasserman Scultz Scott Franklin | X X | X X | |
| GA | Sanford Bishop | X | X | |
| ID | Mike Simpson | X | ||
| IL | Lauren Underwood | X | X | |
| KS | Jerry Moran | X | X | |
| KY | Mitch McConnell | X | X | |
| LA | Julia Letlow Ashley Hinson | X X | X X | |
| MD | Andy Harris Chris Van Hollen | X | X | X |
| ME | Chellie Pingree Susan Collins | X X | X | X |
| MI | John Moolenaar Gary Peters | X X | x | X |
| MN | Betty McCollum | X | X | |
| MS | Cindy Hyde-Smith | X | X | |
| MT | Jon Tester Ryan Zinke | X | X | X |
| NB | Deb Fischer | X | ||
| ND | John Hoeven | X | X | |
| NM | Martin Heinrich | X | X | |
| NV | Mark Amodei | X | ||
| OH | Marcy Kaptur | X | X | |
| OR | Jeff Merkley | X | X | X |
| PA | Guy Reschenthaler | X | X | |
| RI | Jack Reed | X | ||
| TX | Michael Cloud Jake Ellzey | X | X X | |
| UT | Chris Stewart | X | ||
| VA | Ben Cline | X | X | |
| WA | Dan Newhouse Derek Kilmer | X | X X | |
| WV | Shelly Moore Capito Joe Manchin | X | X X | |
| WI | Mark Pocan Tammy Baldwin | X X | X | X |
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice .2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Hudgins, E.J., F.H. Koch, M.J. Ambrose, and B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ 2014; doi:10.1890/130311
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. Frontiers in Ecology.
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Szakonyi, M. 2023. Sourcing shift from China pulls US import share to more than a decade low.