beech leaf disease symptoms; photo courtesy of Jennifer Koch, USFS
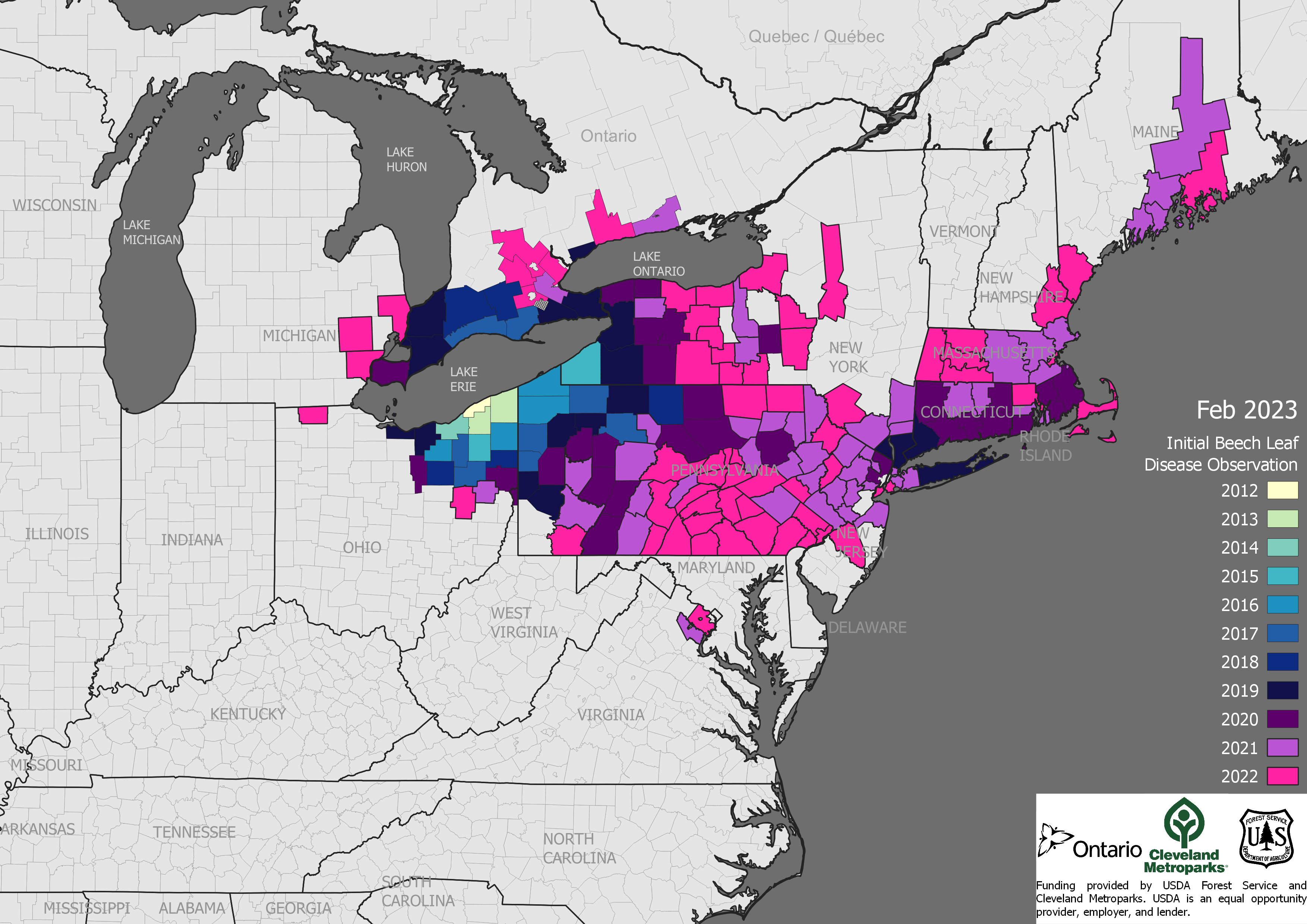
As we know, beech leaf disease (BLD) has spread rapidly in the decade since its discovery in northeast Ohio. It has been detected as far east as the Maine coast, as far south as northern Virginia, as far north as southern Ontario, and as far west as eastern Michigan and northern Indiana. It has been found in 12 states.
BLD is associated with a nematode, Litylenchus crenatae subsp. mccannii (Lcm), although whether this is the sole causal agent is not yet clear.
BLD’s North American host, American beech (Fagus grandifolia),is an important native deciduous hardwood species. It plays important roles in nutrient cycling, erosion control, and carbon storage and sequestration in forests. Wildlife species depend on the trees’ canopies and especially cavities blog for nesting sites, shelter, and nutritious nuts. American beech – with sugar maple (Acer saccharum) and yellow birch (Betula alleghaniensis) – dominate the northern hardwood ecosystem of northeastern United States and southeastern Canada. These forests occupy a huge area; in just New England and New York they occupy 20 million acres (Leak, Yamasaki and Holleran. 2014; full citation at end of blog).
Beech leaf disease also affects European beech, (F. sylvatica), Chinese beech (F. engleriana), and Oriental beech (F. orientalis) planted in North America. The disease has not yet been detected in Asia or Europe. Japanese beech (F. crenata) sporadically display symptomatic leaves, but the disease has not been reported there.
Scientists working to understand the disease, how it spreads, and its ecological impact confer every other month. The next time is in early December.
Paulo Vieira, of the USDA Agriculture Research Service, leads one group seeking to better understand how the disease infects its host. They published a new study (see full citation at end of blog) examining how the nematode provokes changes in the cells of the trees’ leaves. As they point out, leaves are plants’ primary organs for photosynthesis – hence providing energy for growth. The leaf is composed of a several cell types organized into different tissues with specific function related to photosynthesis, gas exchange, and/or the transportation of water and nutrients. Thus, changes in leaf morphology affect the normal functioning of the leaf and therefore the tree’s growth and survival.
Vieira et al. found that:
The BLD nematode enters the leaf bud as it forms in late summer. In early autumn, all nematode developmental stages were found in the buds, including eggs at various stages of embryonic development, juveniles, and adults. Adult males were found in fewer than 20% of the buds, suggesting that the nematode can reproduce asexually.
Feeding by the BLD nematode induces abnormal and extensive cell proliferation, resulting in a significant increase of the number of cell layers inside the leaf. These changes improve the nutrition that the leaves provide to the nematode. However, the BLD-induced distortions of the bud persist as the leaf grows. Symptomatic leaf “banding” results. These areas have a proliferation of abnormally large and irregularly shaped cells with more chloroplasts. Intercellular spaces are also larger; this is where the nematodes are found. in. (The publication has dramatic photographs.)
Sites damaged by nematodes are a major resource for metabolites needed for plant performance. So their damage imposes a considerable drain.
Colonization of roots by ectomycorrhizal fungal is also reduced in severely diseased trees.
Immature female nematodes are the principal winter survivors. However, many die, making it difficult to culture nematodes in the spring. The nematodes reproduce during the growing season. Buildup of nematode numbers makes culturing easier, so facilitating confirmation of the disease’s presence.
Nematodes can migrate along the stem to other leaves, thus spreading the infection.
Vieira et al. tell us fascinating facts about the nematode. The BLD nematode, Litylenchus crenatae subsp. mccannii (Lcm) is now considered one of the top ten most important plant-parasitic nematodes in the United States. To date, species of this genus have been found only in Japan and New Zealand. The species L. crenatae was first described from Japan. A second species — L. coprosma – was detected in 2012 in New Zealand in association with small chlorotic patches on leaves of two native plants in the Coprosma genus.
Litylenchus belongs to the family Anguinidae. Several species in the family are designated quarantine pests because they cause economically significant damage to food and ornamental corps, including grains (wheat, barley, rice) and potatoes. Anguinidae nematodes often parasitize aerial parts of the hosts (e.g., leaves, stems, inflorescences, seeds); less frequently they infest roots. They can migrate along the host tissue surfaces in water films. Their host ranges vary from broad to narrow. Other Anguinidae nematodes apparently share the ability to manipulate the host’s cellular machinery, which often results in the induction of cell hyperplasia [the enlargement of an organ or tissue caused by an increase in the reproduction rate of its cells], and hypertrophy [increase and growth of cells] of the tissues on which they feed.
healthy beech leaves; F.T. Campbell
Vieira et al. assert that the rapid spread of Litylenchus crenatae subsp. mccannii – combined with the apparent lack of resistance in native beech trees – suggests that this nematode was recently introduced to North America. Furthermore, the ability of this subspecies to change the host’s cell cycle machinery supports the link between the presence of the nematode and the disease.
The mechanisms by which nematodes change host-plant cells are unknown. I hope that scientists will pursue these questions. Perhaps the nematode family’s threat to grains and other food crops will prompt funding for such work. Unfortunately, I don’t think the threat to an ecologically-important native tree species will have the same power.
SOURCES
Leak, W.B, M. Yamasaki and R. Holleran. 2014 Silvicultural Guide for Northern Hardwoods in the Northeast. United States Department of Ariculture Forest Service Northern Research Station. General Technical Report NRS-132. April 2014.
Vieira P, M.R. Kantor MR, A. Jansen, Z.A. Handoo, J.D. Eisenback. (2023) Cellular insights of beech leaf disease reveal abnormal ectopic cell division of symptomatic interveinal leaf areas. PLoS ONE October 5, 2023. 18(10) https://doi.org/10.1371/pone.0292588
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
I have been disappointed that a research symposium focused on the northern hardwood forest workshop gave little attention to non-native pests (see citation at end of this blog). A new study based in the Bartlett Experimental Forest in the White Mountains of New Hampshire is more balanced. Ducey et al. (full citation at the end of this blog) analyzed changes in the forest’s species composition and tree size over the past 80 years.
They found that trees of nearly all species are growing into larger sizes as the forest continues to age since the last widespread clearing at the end of the 19th Century. The same aging is causing a rapid decline in two shade-intolerant species – paper birch (Betula papyrifera) and aspen (Populus tremuloides and P. grandidentata) – which had grown quickly once the cleared areas were abandoned. The mid-shade -tolerant species yellow birch (Betula alleghaniensis) also is declining. Together, the birch and aspen species have declined from a quarter to a third of basal area in 1931 to 10 – 12% in 2015.
Some developments are unexpected. Red maple (Acer rubrum) increased in abundance until the early 1990s, but that growth then levelled off. Sugar maple (Acer saccharum) has declined in abundance except where the forest is managed to retain it.
There is little evidence of tree species migrating upward on slopes in response to changes in the local climate. Major weather events – a hurricane in 1938 and an ice storm in 1998 — caused significant tree mortality across Bartlett Experimental Forest, but not a dramatic change in forest composition.
Eastern hemlock (Tsuga canadensis) is replacing the disappearing birch and aspen on low elevation sites. Hemlock has increased its proportion of basal area from 8 – 10% to a quarter or more. Despite aggressive management aimed at reducing the tree’s presence, American beech (Fagus grandifolia) is on track to dominate large areas of the Bartlett Experimental Forest. Given the tree-killing pests already present in the region, large increases in eastern hemlock, American beech, and red spruce (Picea rubens) are worrying.
Eastern hemlock creates important wildlife habitat for deer and more than 100 other vertebrate species in New England. However, hemlock woolly adelgid (HWA) has been present in New Hampshire since 2000. It is now within 15-20 km of Bartlett Experimental Forest. There is some hope that the region’s cold temperatures might limit HWA’s spread and impacts, but Ducey et al. expect major change when the adelgid arrives.
beech saplings; photo by FT Campbell
Ducey et al. cite a separate study demonstrating that mortality caused by beech bark disease (BBD) can be sufficient to upset carbon storage in old-growth forests. On the Bartlett Forest, nearly 90% of beech trees had become diseased by 1950.
Ducey et al. express concern about the possible impact of beech leaf disease (BLD), as well.
BLD has not yet been detected in the White Mountains or New Hampshire, but is in so New England and coastal Maine. Much remains unknown about the disease, including how it spreads and its long-term impacts.
Ducey et al. do not raise pest concerns about red spruce or balsam fir (Abies balsamea), which co-dominate the Bartlett Forest at higher elevations (above 500 m). This silence is disturbing since red spruce can be killed by the brown spruce longhorned beetle, a European woodborer established in Nova Scotia and threatening to spread south. Balsam firs suffer some mortality from feeding by the balsam woolly adelgid, a Eurasian sap-sucker which has been in New England for more than a century.
brown spruce longhorned beetle
White ash (Fraxinus americana) is present as a minor component of the Bartlett Forest. Because it is considered to be a valuable timber species, management has resulted in a modest increase in abundance of ash. Ducey et al. expect dramatic reduction — or even elimination of the species — when the emerald ash borer (EAB) arrives. EAB has been detected within ~ 15 km from Bartlett Experimental Forest.
Ducey et al. conclude that silvicultural management applied at the scope and intensity of that in the Bartlett Experimental Forest has moderated some changes. That is, it is maintaining sugar maple and suppressing the increase of beech. Its effect is secondary, however to overall forest development as the forest ages.
SOURCES
Ducey, M.J, O.L., Yamasaki, M. Belair, E.P., Leak, W.B. 2023. Eight decades of compositional change in a managed northern hardwood landscape. Forest Ecosystems 10 (2023) 100121
Proceedings of the First Biennial Northern Hardwood Conference 2021: Bridging Science and Management for the Future. USDA Forest Service Northern Research Station General Technical Report NRS-P-211, May 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
tanoak trees in southern Oregon killed by sudden oak death; photo by Oregon Department of Forestry; this pathogen is not mentioned by USFS RPA report
In August the USDA Forest Service published the agency’s 2020 assessment of the future of America’s forests under the auspices of the Resources Planning Act. [See United States Department of Agriculture Forest Service Future of America’s Forests and Rangelands, full citation at the end of the blog.] To my amazement, this report is the first in the series (which are published every ten years) to address disturbance agents, specifically invasive species. In 2023! Worse, I think its coverage of the threat does not reflect the true state of affairs – as documented by Forest Service scientists among others.
This is most unfortunate because policy-makers presumably rely on this report when considering which threats to focus on.
Here I discuss some of the USFS RPA report and what other authors say about the same topics.
The RPA Report’s Principle Foci: Extent of the Forest and Carbon Sequestration
The USFS RPA report informs us that America’s forested area will probably decrease 1- 2% over the next 50 years (from 635.3 million acres to between 619 and 627 million acres), due largely to conversion to other uses. This decline in extent, plus trees’ aging and increases in disturbance will result in a slow-down in carbon sequestration by forests. In fact, if demand for wood products is high, or land conversion to other uses proceeds apace, U.S. forest ecosystems are projected to become a net source of atmospheric CO2 by 2070.
Eastern forests sequester the majority of U.S. forest carbon stocks. These forests are expected to continue aging – thereby increasing their carbon storage. Yet we know that these forests have suffered the greatest impact from non-native pests.
I don’t understand why the USFS RPA report does not explicitly address the implications of non-native pests. In 2019, Songlin Fei and three USFS research scientists did address this topic. Fei et al. estimated that tree mortality due to the 15 most damaging introduced pest species have resulted in releases of an additional 5.53 terragrams of carbon per year. Fei and colleagues conceded this is probably an underestimate. They say that annual levels of biomass loss are virtually certain to increase because current pests are still spreading to new host ranges (as demonstrated by detection of the emerald ash borer in Oregon). Also, infestations in already-invaded ranges will intensify, and additional pests will be introduced (for example, beech leaf disease).
I see this importance of eastern forests in sequestering carbon as one more reason to expand efforts to protect them from new pest introductions, and the spread of those already in the country, etc.
A second issue is the role of non-native tree species in supporting the structure and ecological functions of forests. Ariel Lugo and colleagues report that 18.8 million acres (7.6 million ha, or 2.8% of the forest area in the continental U.S.) is occupied by non-native tree species. (I know of no overall estimate for all invasive plants.) They found that non-native tree species constitute 12–23% (!) of the basal area of those forest stands in which they occur.
Norway maple (Acer platanoides); one of the most widespread invasive species in the East. Photo by Hermann Falkner via Flickr
Lugo and colleagues confine their analysis of ecosystem impacts to carbon sequestration. They found that the contribution of non-native trees to carbon storage is not significant at the national level. In the forests of the continental states (lower 48 states), these trees provide 10% of the total carbon storage in the forest plots where they occur. (While Lugo and colleagues state that the proportion of live tree biomass made up of non-native tree species varies greatly among ecological subregions, they do not provide examples of areas on the continent where their biomass – and contribution to carbon storage — is greater than this average.) In contrast, on Hawai`i, non-native tree species provide an estimated 29% of live tree carbon storage. On Puerto Rico, they provide an even higher proportion: 36%.
Brazilian pepper (Schinus terebinthifolius) – widespread invasive in Hawai`i and Florida; early stage invasive in Puerto Rico. Photo by Javier Alexandro via Flickr
In the future, non-native trees will play an even bigger role. Since tree invasions on the continent are expanding at ~500,000 acres (202,343 ha) per year, it is not surprising that non-native species’ saplings provide 19% of the total carbon storage for that size of trees in the lower 48 states (Lugo et al.).
Forming a More Complete Picture: Biodiversity, Disturbance, and Combining Data.
The USFS RPA report has a chapter on biodiversity. However, the chapter does not discuss historic or future diversity of tree species within biomes, nor the genetic diversity within tree species.
Treatment of Invasive Species
The USFS 2020 RPA report is the first to include a chapter on disturbance, including invasive species. I applaud its inclusion while wondering why they have included it only now? Why is the coverage so minimal? I think these lapses undercut the report’s purpose. The RPA is supposed to inform decision-makers and stakeholders about the status, trends, and projected future of renewable natural resources and related economic sectors for which USFS has management responsibilities. These include: forests, forest products, rangelands, water, biological diversity, and outdoor recreation. The report also has not met its claim to “capitalize on” areas where the USFS has research capacity. One excuse might be that several important publications have appeared after the cut-off date for the assessment (2020). Still, the report’s authors cite some of the evaluations that were in preparation as of 2020, e.g., Poland et al.
I suggest also that it would be helpful to integrate data from other agencies, especially the invasive species database compiled by the U.S. Geological Survey, into the RPA. For example, the USGS lists just over 4,000 non-native plant species in the continental U.S. (defined as the lower 48 plus Alaska). On Hawai`i, the USGS lists 530 non-native plant species as widespread. Caveat: many of the species included in these lists probably coexist with the native plants and make up minor components of the plant community.
Specifically: Invading Plants
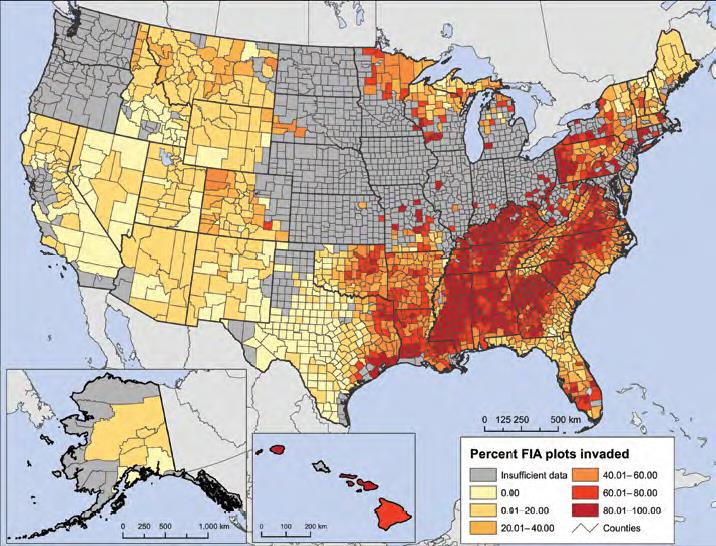
The USFS RPA report gives much more attention to invasive plants than non-native insects and pathogens. The report relies on the findings of Oswalt et al., who based their data on forested plots sampled by the Forest Inventory and Analysis (FIA) program. (The RPA also reports on invasive plants detected on rangelands, primarily grasslands.) Oswalt et al. found that 39% of FIA plots nationwide contained at least one plant species that the FIA protocol considers to be invasive and monitors. The highest intensity of plant invasions is in Hawai`i – 70% of the plots are invaded. The second-greatest intensity is in the eastern forests: 46%. However, the map showing which plots were inventoried for invasive plants makes clear how incomplete these data are – a situation I had not realized previously.
I appreciate that the USFS RPA report mentions that propagule pressure is an important factor in plant invasions. This aspect has often been left out in past analyses. I also appreciate the statement that international trade in plants for ornamental horticulture will probably lead to additional introductions in the future. Third, I concur with the report’s conclusions that once forest land is invaded, it is unlikely to become un-invaded. Invasive plant management in forests often results in one non-native species being replaced by another. In sum, the report envisions a future in which plant invasion rates are likely to increase on forest land.
If you wish to learn more about invasive plant presence and impacts, see the discussion of invasive plants in Poland et al., my blogs based on the work by Doug Tallamy, and several other of my blogs compiled under the category “invasive plants” on this website.
I believe all sources expect that the area invaded by non-native plant species, and the intensity of existing invasions, will increase in the future.
The USFS RPA links these invasions to expansion of the “wildland-urban interface” (“WUI”). These areas increased rapidly before 2010. At that time, they occupied 14% of forest land. The report published in 2023 did not assess their future expansion over the period 2020 to 2070. However, it did project increased fragmentation in many regions, especially in the RPA Western and Southeastern regions. Since “fragmentation” is very similar to wildland-urban interfaces, the report seems implicitly to project more widespread plant invasions in the future.
plant invasions facilitated by fragmentation; northern Virginia; photo by F.T. Campbell
Specifically: Insects and Pathogens
The USFS RPA report on insects and pathogens is brief and contains puzzling errors and gaps. It says that the tree canopy area affected by both native and non-native mortality-causing agents has been consistently large over the three most recent five-year FIA assessment periods. It notes that individual insects or diseases have extirpated entire tree species or genera and fundamentally altered forests across broad regions. Examples cited are chestnut blight and emerald ash borer.
The USFS RPA report warns that pest-related mortality might be underreported in the South, masked by more intense management cycles and higher rates of tree growth and decay. On the other hand, the report asserts that pest-related mortality is probably overrepresented in the Northern Region in the 2002 – 2006 period because surveyors drew polygons to encompass large areas affected by EAB and balsam woolly adelgid (Adelges piceae) infestations. The latter puzzles me; I think it is probably an error, and should have referred to hemlock woolly adegid, A. tsugae. Documented mortality has generally been much more widespread from insects than diseases, e.g., bark beetles, including several native ones, across all regions and over time, especially in the West – where the most significant morality agents are several native beetles. The USFS RPA report mentions that the Northern Region has been particularly affected by non-native pests, including EAB, HWA, BWA, beech bark disease, and oak wilt. It mentions that Hawai`i has also suffered substantial impacts from rapid ʻōhiʻa death.
Defoliating insects have affected relatively consistent area over time. This area usually equaled or exceeded the area affected by the mortality agents. Principal non-native defoliators in the Northern Region have been the spongy moth (Lymantria dispar); larch casebearer (Coleophora laricella); and winter moth (Operophtera brumata). In the South they list the spongy moth.
More disturbing to me is the USFS RPA report’s conclusion that the future impact of forest insects is highly uncertain. The authorsblame the complexity of interactions among changing climate, those changes’ effects on insect and tree species’ distributions, and overall forest health. Also, they name uncertainty about which new non-native species will be introduced to the United States. I appreciate the report’s avoidance of blanket statements regarding the effects of climate change. However, other studies – e.g., Poland et al. – have incorporated these complexities while still offering conclusions about a number of currently established non-native pests. Finally, I am particularly dismayed that the USFS RPA does not provide analysis of any forest pathogens beyond the single mention of a few.
I am confused as to why the USFS RPA report makes no mention of Project CAPTURE (Conservation Assessment and Prioritization of Forest Trees Under Risk of Extirpation). This is a multi-partner effort to prioritize U.S. tree species for conservation actions based on invasive pests’ threats and the trees’ ability to adapt to them. Several USFS units participated, including the Southern Research Station, the Eastern Forest Environmental Threat Assessment Center, and the Forest Health Protection program. The findings were published in 2019. See here. Lead scientist Kevin Potter was one of the authors of the RPA’s chapter on disturbance.
redbay (Persea borbonia) trees in Georgia killed by laurel wilt; photo by Scott Cameron. Redbay is ranked by Project CAPTURE as 5th most severely at risk due to a non-native pest
“Project CAPTURE” provided useful summaries of non-native pests’ impacts, including the facts that
54% of the tree species on the continent are infested by one or more non-native insect or pathogen;
nearly 70% of the host/agent combinations involve angiosperm (broadleaf) species, 30% gymnosperms (e.g., conifers). When considering only non-native pests, pests attacking angiosperms had greater average severity.
Disease impacts are more severe, on average, than insect pests. Wood-borers are more damaging than other types of insect pests.
Non-native agents have, on average, considerably more severe impacts than native pests.
Project CAPTURE also ranked priority tree species based on the threat from non-native pests (Potter et al., 2019). Tree families at the highest risk to non-native pests are: a) Fagaceae (oaks, tanoaks, chestnuts, beech), b) Sapindaceae (soapberry family; includes maples, Aesculus (buckeye, horsechestnut); c) in some cases, Pinaceae (pines); d) Salicaceae (willows, poplars, aspens); e) Ulmaceae (elms) and f) Oleaceae (includes Fraxinus). I believe this information should have been included in the Resources Planning Act report in order to insure that decision-makers consider these threats in guiding USFS programs.
I also wish the USFS RPA had at least prominently referred readers to Poland et al. Among that study’s key points are:
Invasive (non-native) insects and diseases can reduce productivity of desired species, interactions at other trophic levels, and watershed hydrology. They also impose enormously high management costs.
Some non-native pests potentially threaten the survival of entire tree genera, not just individual species, e.g., emerald ash borer and Dutch elm disease. I add white pine blister rust and laurel wilt.
Emerald ash borer and hemlock woolly adelgid are listed as among the most significant threats to forests in the Eastern US.
White pine blister rust and hemlock woolly adelgid are described as so profoundly affecting ecosystem structure and function as to cause an irreversible change of ecological state.
Restoration of severely impacted forests requires first, controlling the non-native pest, then identifying and enriching – through selection and breeding – levels of genetic resistance in native populations of the impacted host tree. Programs of varying length and success target five-needle pines killed by Cronartium ribicola; Port-Orford cedar killed by the oomycete Phytophthora lateralis; chestnut blight; Dutch elm disease; butternut canker (causal agent Ophiognomonia clavigignenti juglandacearum), emerald ash borer; and hemlock woolly adelgid.
Climate change will almost certainly lead to changes in the distribution of invasive species, as their populations respond to increased variability and longer-term changes in temperature, moisture, and biotic interactions. Predicting how particular species will respond is difficult but essential to developing effective prevention, control, and restoration strategies.
Poland et al. summarizes major bioinvaders in several regions. Each region except Hawai`i (!!) includes tree-killing insects or pathogens.
It is easier to understand the RPA report’s not mentioning priority-setting efforts by two other entities, the Morton Arboretumand International Union for the Conservation of Nature (IUCN). These studies were published in 2021 and their lead entities were not the Forest Service – although the USFS helped to fund the U.S. portion of the studies.
The Morton Arboretum led in the analysis of U.S. tree species. It published studies evaluating the status of tree species belonging to nine genera, considering all threats. The Morton study ranked as of conservation concern one third of native pine species; 31% of native oak species; significant proportion of species in the Lauraceae. The report on American beech — the only North American species in the genus Fagus – made no mention of beech leaf disease – despite it being a major concern in Ohio – only two states away from the location of the Morton Arboretum near Chicago.
valley oak (Quercus lobata) in Alameda Co, California; photo by Belinda Lo via Flickr
Most of the species listed by the Morton Arboretum are of conservation concern because of their small populations and restricted ranges. The report’s coverage of native pests is inconsistent, spotty, and sometimes focuses on odd examples.
Tree Species’ Regeneration
Too late for consideration by the authors of the USFS RPA report come new studies by Potter and Riitters that evaluate species at risk due to poor regeneration. This effort evaluated 280 forest tree species native to the continental United States – two-thirds of the species evaluated in the Kevin Potter’s earlier analysis of pest impacts.
The results of Potter and Riitters 2023 only partially matched those of the IUCN/Morton studies. The Morton study did not mention three genera with the highest proportions of poorly reproducing species according to Potter and Riitters: Platanus,Nyssa, and Juniperus. Potter, Morton, and the IUCN largely agree on the proportion of Pinus species at risk. Potter et al. 2023 found about 11% of oak species to be reproducing poorly, while Morton designated a third of 91 oak species to be of conservation concern.
I believe Potter and Riitters and the Morton study agree that the Southeast and California are geographic hot spots of tree species at risk.
Potter and Riiters found that several species with wide distributions might be at risk because they are reproducing at inadequate rates. Three of these exhibit poor reproduction across their full range: Populus deltoids (eastern cottonwood), Platanus occidentalis (American sycamore), and ponderosa pine(Pinus ponderosa). Four more species are reported to exhibit poor reproduction rates in all seed zones in which they grow (the difference from the former group is not explained). These are two Juniperus,Pinus pungens, and Quercus lobata. As I point out in my earlier blog, valley oak is also under attack by the Mediterranean oak borer.
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National Academy of Sciences. Vol. 116, No. 35. August 27, 2019.
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, and C.M. Kurtz. 2022. The Contribution of Nonnative Tree Species to the Structure and Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific and Caribbean. USDA USFS General Technical Report IITF-54
Poland, T.M., T. Patel-Weynand, D.M. Finch, C.F. Miniat, D.C. Hayes, V.M. Lopez, eds. 2021. Invasive Species in Forests and Rangelands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. Available gratis at https://link.springer.com/book/10.1007/978-3-030-45367-1
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation.
Potter, K.M. and Riitters, K. 2023. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests 2022, 13, 19. https://doi.org/10.3390/f13010019
United States Department of Agriculture Forest Service. 2023. Future of America’s Forests and Rangelands: The Forest Service 2020 Resource Planning Act Assessment. GTR-WO-102 July 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Eastern deciduous forest is large and important ecologically. The forest is important for biological diversity: it shelters many endangered species, especially plants, molluscs and fish, mammals, and reptiles. In addition, the majority of forest carbon stocks in the U.S. are those of theeastern states.
But the Eastern deciduous forest is also under many anthropogenic stresses – including high numbers of non-native insects and pathogens, Liebhold map high numbers of invasive plants, blog invasive earthworms, blog browsing by overabundant deer, and timber extraction. In the southern portions of the forest, human populations are expanding, resulting in landscape fragmentation (USDA FS 2023b RPA, full reference at end of this blog).
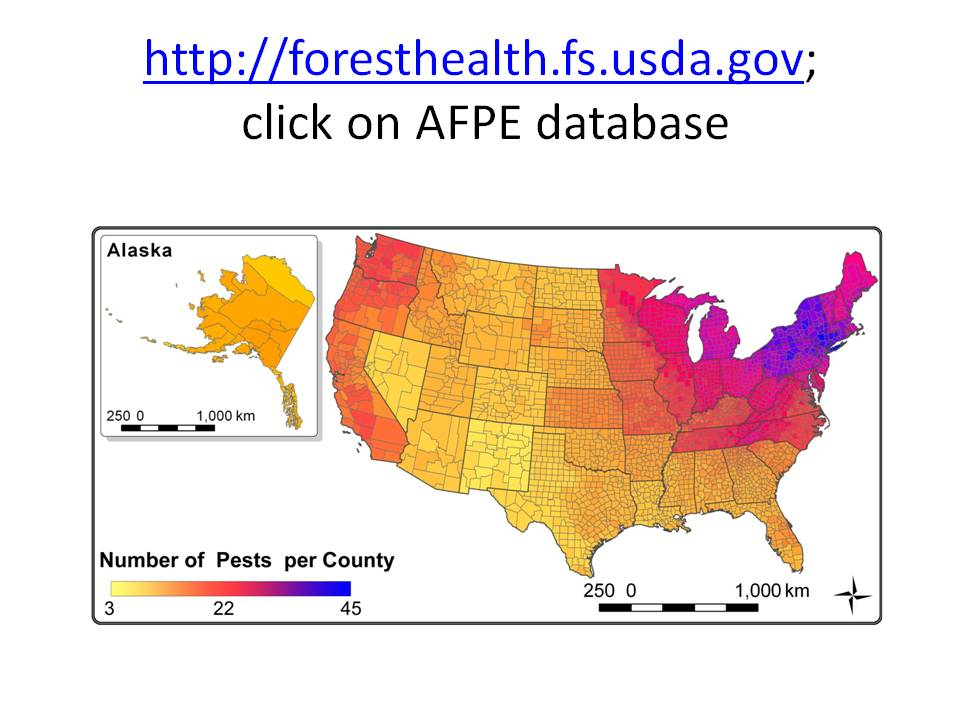
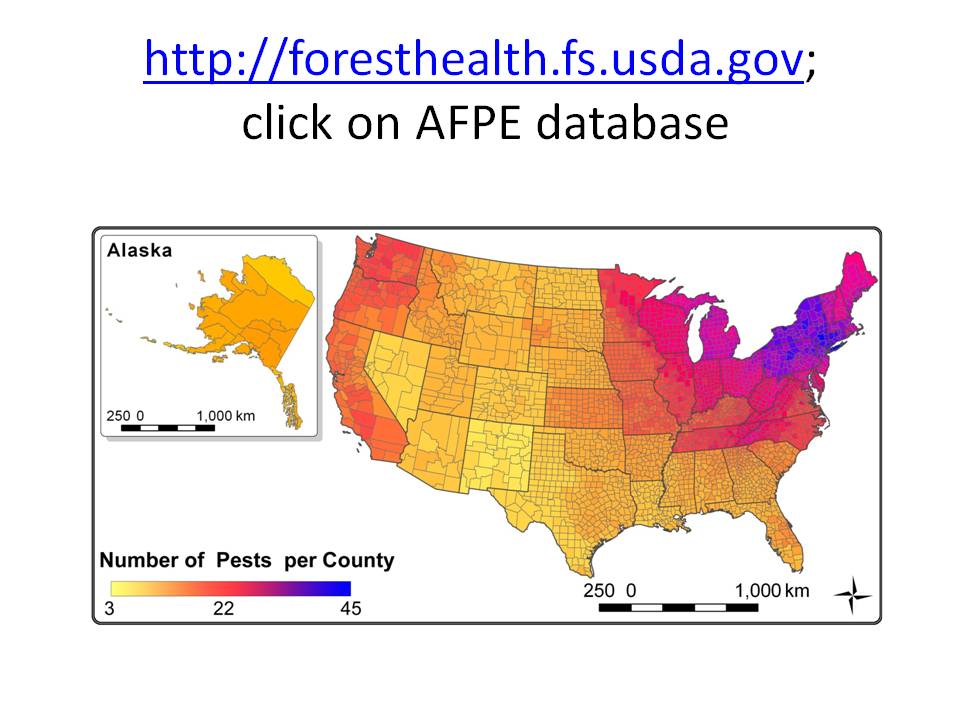
map showing number of non-native pests in each county, as of ~2010
Agency and academic scientists in the USDA Forest Service Eastern Region (Maine to Minnesota; Delaware to West Virginia, then north of the Ohio River to Missouri) are trying to understand how long-term, continuous stressors, like deer browsing and invasive plants and earthworms, – interact with short-term gap-forming events. They call the long-term stressors “press” disturbances to distinguish them from the short-term “pulse” disturbances (Reed, Bronson, et al.; full citation at end of this blog). Understanding the processes by which forests recover from disturbance is increasingly important. Climate change is expected to raise the frequency and intensity of catastrophic natural disturbances (Spicer and Reed, Royo et al.).
The scientists emphasize that the impacts of these stressors – and effective solutions — vary depending on context.
Invasive Earthworms
USDA APHIS is responsible for regulating introduction of new species. For earthworms, APHIS’ principal concern is clearly the possibility that imported worms or soil might transport pathogens. However, the agency’s website does mention worms’ ability to disrupt the soil and possibly cause undesirable impacts on plant growth and diversity. At the 2023 National Plant Board meeting in early August 2023, Gregg Goodman, Senior Agriculturalist in APHIS PPQ NPB website for agenda? discussed issues that he considers when evaluating whether to grant permits for importing earthworms. APHIS allows imports to be used for fish bait. Dr. Goodman explained that APHIS surveyed fishermen to determine where they dump unused bait. He found no damage to plants along streams, etc. where they are dumped. A state plant health official from a northern state and I objected that the ecosystem damage caused by earthworms is well documented and we doubted that dumping of bait is not a pathway for introducing worms into natural areas.
Reed, Bronson et al. found lower earthworm biomass and density in both deer exclosures and canopy gaps. They hypothesize that the new plant growth associated with canopy gaps attracts deer, resulting in increased browse pressure. That browse pressure then affects the plant community, succession and forest structure. The changed plant community affects soil properties that then affect soil-dwelling fauna like earthworms. They believe the higher worm densities in closed-canopy sites might be the result of nutrient-rich tree leaf litter which provides both shelter and food. Another factor might be lack of recent soil disturbances in closed canopy sites.
While they say need more research is needed on the complex, combined effects of earthworms and deer, Reed, Bronson et al. still suggest that reducing deer populations or – where that is not possible – creating gaps might help manage earthworm invasions.
Deer Interactions
The long-term, chronic effect of excessive deer herbivory are well documented. See the many presentations at the recent Northern Hardwood research forum (USDA FS 2023b Proceedings). Most studies show that deer browsing overwhelms other disturbances, such as fire and canopy gaps that typically promote seedling diversity. However, recent results refine our understanding.
Samuel P. Reed and colleagues (Reed, Royo et al.) found that on the Allegheny Plateau of western Pennsylvania high deer densities at the time of stand initiation resulted in long-term reduced tree species diversity, density, and basal area. These responses were still detectable nearly four decades later. Stands are dominated by the unpalatable black cherry (Prunus serotina). The reduced stand density and the cherries’ narrower crowns lead to less above-ground biomass and reductions in above-ground carbon stocks. These scientists recommend that managers reduce deer populations to prevent changes in forest structure with probably long-term and important ramifications for many ecosystem functions.
Prunus serotina; photo by Awinch1001 via Flickr
Hoven et al. concurred with the importance of reducing deer densities, but suggested focussing on wet sites where, in their study, deer browsing had its greatest effects. On drier sites deer browsing had no effect on the diversity of woody plant seedlings.
Spicer et al. seek particularly to maintain a heterogeneous landscape to allow coexistence of both early- and late-successional species. In the Eastern Deciduous Forest biome, herbs, shrubs, and vines comprise 93% of the species richness of vascular plants
These authors found that the impact of deer browsing diverged depending on vegetation management actions. In wind-throw gaps where the plant community was retained, deer caused a 14% decline in shrub cover. In contrast, when scientists removed the extant vegetation at the beginning of recovery, deer exclusion caused a 67% increase in shrub cover. The authors speculate that vegetation removal stimulated abundant blackberry (Rubus species) regrowth. Where they had access (in gaps lacking exclosures), deer heavily browsed young Rubus stalks that sprouted after the competing vegetation was cut down. However, when the pre-established vegetation was not removed, older Rubus thickets might have protected other herbs and shrubs from browsing. Spicer et al. did not observe any major shifts in browse-tolerant species in deer-exclusion plots.
Invasive Shrubs
Hoven et al. found that in drier forest plots, the presence of non-native shrubs reduced native seedling abundance, richness, and diversity. Instead there were more seedlings of introduced species, including Lonicera maackii, L. morrowii, Ligustrum sp., and Rosa multiflora. They are concerned that replacement by invasive honeysuckles might be particularly strong in gaps resulting from death of ash trees caused by emerald ash borer. Woodlands could become dominated introduced shrubs, reducing diversity. Consequently, they recommend removing non-native shrubs in drier forests to promote seedling numbers and diversity.
In contrast, in wetter forests basal area of non-native shrubs did not affect introduced seedling abundance. However, the shrubs’ size did promote greater proportions of Lonicera maackii and Ligustrum seedlings. They suggest this might be the outcome of either abundant seed sources or allelopathic properties of some invasive shrubs e.g., L. maackii. In such sites, seedling diversity is already limited to plants that tolerate waterlogging. A hopeful note is that one native shrub, Lindera benzoin, seems able to prevent establishment of L. maackii.
Lonicera maackii; photo by pverdonk via Flickr
Hoven et al. do worry that death of ash trees might lead to declining transpiration rates, raising water tables, and further reducing seedling species richness and diversity.
Impact of Salvage Logging and Vegetation Removal
Spicer et al. studied how anthropogenic stressors affect succession. These scientists took advantage of tornado-caused gaps to compare interactions with deer browsing, salvage logging, and mechanical removal of the understory.
Contrary to expectations, none of these anthropogenic disturbances delayed community recovery or reduced diversity in comparison to the natural disturbance (tornado blowdown). Instead, adding either salvage logging or mechanical removal of understory vegetation substantially enhanced herbaceous species richness and shrub cover.
However, each major plant growth form responded differently. First, none of the manipulations affected species diversity or abundance of tree seedlings and saplings. Second, salvage logging in the wind-throw gaps increased species richness of herbs by 30%. Shrub abundance was doubled and cover almost tripled, but species richness did not change. Third, removing competing understory vegetation caused an increase of 23% in mean herbaceous cover. I have already discussed the impact of excluding deer.
Spicer et al. greet these increases in species richness with enthusiasm; they recommend managing to create a patchwork of combined natural and anthropogenic disturbances to promote plant diversity. However, I have some questions about which species are being promoted.
This study identified a total of 264 vascular plant species: 40 trees, 190 herbs, 15 shrubs, 17 vines, and 2 of unknown growth form. Only about half of these, 123 species, grew in portions of the mature forest not affected by either the tornado or one of the anthropogenic manipulations.
Gaps contained more plant species – as is to be expected. Natural blowdown areas where no manipulation was carried out had 49 more species than the undisturbed forest community (172 species). Blowdown sites subjected to salvage logging added another 53 species for a total of 225 species, or 102 more than the undisturbed reference forest.
A total of 17 species occurred only once in the authors’ data [= unique species]. Eight of these species grew only in the undisturbed forest. Two grew only in the tornado-impacted plots. Spicer et al. do not elaborate on whether these species are officially rare in that part of Pennsylvania – although it seems they might be. I wish Spicer et al. had addressed whether these possibly rare species might be affected by the forest management they recommend, i.e., intentionally creating a patchwork of various disturbances. An additional seven unique species were found in plots that had been subjected to an anthropogenic disturbance — either salvage logging or removal of remnant vegetation.
nodding trillium (Trillium cernuum); imperiled by restricted range or low populations; photo by Jason Ryndock, Pennsylvania Natural Heritage Program
In tornado-disturbed sites, one native species associated with areas where vegetation was left intact is one of the gorgeous wildflowers of eastern deciduous forests: a Trillium (species not indicated). The one native plant associated with plots from which vegetation was removed was a grass (unspecified).
Spicer et al. report that the proportion of the flora composed of non-native species was very similar between the salvage-logged area (7%) and the undisturbed reference forest (5%). Half of the non-native plant species (3.5% or 9 species) are listed as invasive in Pennsylvania (the article does not list them).
Spicer et al. say these non-native species are relatively uncommon and that they pose a minimal threat. They do concede that the invasive thorny shrub barberry (Berberis thunbergii) was more common in disturbed than intact areas. [I saw plenty of barberry along forest edges in Cook State Forest, which is only 100 miles away from the study site.] I think Spicer and others are too blasé since invasive plant populations can build up quickly when seed sources are present.
Spicer et al. raise two caveats. First, their results regarding the beneficial effects of salvage logging and vegetation manipulation probably will not apply to situations in which vast areas are logged.
More pertinent to us, they warn that their results would also not apply to forest areas in which propagules have been drastically depleted. This can result from previous human land-use or repeated catastrophic disturbances, such as canopy fires. Nor would their results apply to forests that are more threatened by invasive species. They note that a widespread and dense understory of multiple non-native species can create invasional meltdowns, resulting in a lasting depauperate state. This is especially the case when invaders at higher trophic levels, such as earthworms, are part of the mix.
Other Lessons
Reed, Bronson et al. conclude that forest canopies’ responses to disturbance are too variable to be measured by a single method. Evaluating proposals for management will require multiple measures. The overwhelming recommendation of presenters at the recent northern hardwoods research symposium (USDA FS 2023a Proceedings) was to adapt more flexible management strategies to promote forest sustainability and species diversity.
Hovena et al.’s principal finding is that interactions among site wetness, non-native shrubs and the total basal area of trees in the stand had the largest impacts on the species composition of seedlings. In Ohio, site wetness and chronic stressors like deer and introduced shrubs are acting together to shift seedling communities towards fewer native species. Of these three long-term “press” stresses, the interaction between introduced shrubs and soil wetness overshadowed even the impact of deer herbivory on seedling species richness and abundance. Surprisingly, site-specific characteristics – e.g., wetness, canopy tree competition, deer herbivory and introduced shrubs – were more influential than ash mortality in shaping woody seedling communities.
SOURCES
Hoven, B.M., K.S. Knight, V.E. Peters, D.L. Gorchov. 2022. Woody seedling community responses to deer herbivory, introduced shrubs, and ash mortality depend on canopy competition and site wetness. Forest Ecology and Management 523 (2022) 120488
Reed, S.P., D.R. Bronson, J.A. Forrester, L.M. Prudent, A.M. Yang, A.M. Yantes, P.B. Reich, and L.E. Frelich. 2023. Linked disturbance in the temperate forest: Earthworms, deer, and canopy gaps. Ecology. 2023;104:e4040. https://onlinelibrary.wiley.com/r/ecy
Reed, S.P, A.A. Royo, A.T. Fotis, K.S. Knight, C.E. Flower, and P.S. Curtis. 2022. The long-term impacts of deer herbivory in determining temperate forest stand and canopy structural complexity. Journal of Applied Ecology. 2022; 59:812-821
Spicer, M.E., A.A. Royo, J.W. Wenzel, and W.P. Carson. 2023. Understory plant growth forms respond independently to combined natural and anthropogenic disturbances. Forest Ecology and Management 543 (2023) 12077
United States Department of Agriculture. Forest Service. 2023a. Proceedings of the First Biennial Northern Hardwood Conference 2021: Bridging Science and Management for the Future. Northern Research Station General Technical Report NRS-P-211 May 2023
United States Department of Agriculture. Forest Service. 2023b. Future of America’s Forests and Rangelands. Forest Service 2020 Resources Planning Act Assessment. GTR-WO-102. July 2023 https://www.fs.usda.gov/research/treesearch/66413
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
At CISP, our hearts go out to all those affected by the terrible August fires on Maui. May the departed rest in peace. May the living find comfort and all that is needed for recovery.
Fire and Invasive Grasses
A fire in non-native grasses on Maui in 2009; photo by Forrest and Kim Starr
Major U.S. and international media continue to detail the fires’ devastation, especially in Lahaina. As time has passed, more news has highlighted the role that the widespread presence of introduced, fire-prone grasses played in the rapid growth and spread of Maui’s fires.
For example, The Washington Post devoted seven paragraphs in one story to the issue of grasses. The story quotes several experts: Alison Nugent, an associate atmospheric scientist at the University of Hawaii’s Water Resources Research Center; Jeff Masters, a meteorologist for Yale Climate Connections; and Clay Trauernicht, a fire researcher at the University of Hawaii.
These and others have been widely quoted in the many recent articles. I am glad that they – and the media – are making clear that climate change is not the sole factor causing damaging wildfires. It is clear that Maui’s recent weather patterns – including the high-velocity winds and drought – have been within the range of normal climate patterns. Fluctuations in the Pacific’s weather have also been normal, especially under the influence of the current El Niño.
The dangers caused by Hawai’i’s fire-prone grasses are also clear – and have been for years. Experts have identified policy weaknesses at the county and state level. Also, they have specified changes to land management that could better prevent or mitigate wildfires. There has been far too little action.
On the other hand, there are hopeful signs.
endangered ‘akikiki photo by Carter Atkinson, USGS
The Hawai’i Wildfire Management Organization, a nonprofit, is educating and engaging communities state-wide. Elizabeth Pickett, a Co-Executive Director, presented an overview of wildfire at the Hawai’i Invasive Species Awareness Month in February 2023. The Big Island Invasive Species Committee has successfully eradicated two species of pampas grass on Hawai’i Island – after 13 years’ work. A native species has been planted where pampas formerly grew.
Another Postarticle reported on efforts by staff and fire departments to protect the Maui Bird Conservation Center, which houses critically endangered Hawaiian birds found nowhere else on Earth, including some currently extinct in the wild. As I have blogged previously, the palila, kiwikiu, ‘akikiki, ‘alalā [Hawaiian crow; extinct in the wild] and other birds are dying from avian malaria, carried by nonnative mosquitoes. The Center on Maui and another on the Big Island are run by the San Diego Zoo Wildlife Alliance. Conservationists have completed field trials of a proposed mosquito suppression process for Maui and are seeking public comments for a similar program on Kaua’i. These programs represent groundbreaking and long-awaited progress on countering a principal threat to the survival of Hawai`i’s unique avifauna.Loss of the Center and its birds would have devastated post-suppression efforts to rebuild and restore bird populations in the wild.
The Post carried a second story about the effort to protect Hawai`i’s endangered birds – a full page of print, even longer – with many photos, on the web. The article mentions the “Birds, Not Mosquitoes” program and varying views about it. I rejoice that the dire situation for the Islands’ biodiversity is getting attention in the Nation’s capital. Again, see my earlier blog.
Plant Invasions in Hawaiian Forests
A team of scientists from the USDA Forest Service and Natural Resources Conservation Service, plus the Hawaii Division of Forestry and Wildlife, has carried out a new assessment of the extent of invasive plant species in forests on the Hawaiian Islands (Potter et al. 2023; full citation at end of blog).
The results of their analysis are – in their words – “sobering”. They portend “a more dire future for Hawai`i`s native forests.”
First, regarding the recent fires, Potter et al. found significantly higher cover by invasive grasses on Forest and Inventory Analysis (FIA) plots on Hawai‘i and Maui than on O‘ahu, Kaua‘i, and Lana‘i. Grass invasions were particularly high on the eastern coast of Maui – near Lahaina. Even so, the authors say their study’s methods resulted in a gross underestimate of areas invaded by fire-prone grasses. That is, most of Hawai’i’s xerophytic dry forests were converted to grasslands before the FIA program began. Therefore these grasslands are not included in FIA surveys.
Psidium cattleyanum; photo by Forrest and Kim Starr
The extent of current invasions in wetter forests is already significant – but trends point to an even more worrying future.
Naturalized non-native plant taxa constitute half of the Hawaiian flora.
56% of Hawaii’s 553,000 ha of forest land contained non-native tree species; about 39% of these forest lands are dominated by non-native tree species. Invasive plant species of particular concern were found in the understory of 27% of surveyed forest plots.
Across all islands, six of the ten most abundant species are non-native: Psidium cattleyanum,Schinus terebinthifolius, Leucaena leucocepahala, Ardisia elliptica, Psidium guajava, and Acacia confusa.
While less than one-third (29%) of large trees across the Islands are non-native, this proportion increases to about two-thirds of saplings (63%) and seedlings (66%). Potter et al. focus on the likelihood that plant succession will result in transformation of these forests’ canopies from native tree species to non-native species.
75% of forests in lower-elevation areas of all islands are already dominated by non-native tree species. “Only” 31% of higher-elevation forests are so dominated. These montane forests have been viewed as refugia for native species, but all are invaded to some extent – and likely to become more degraded.
Potter et al. say the high elevation forests might be more resistant to domination by non-natives. Such a result would be counter to well-documented experience, though. Even the authors report that the montane rainforests and mesophytic forests of O‘ahu and Kaua‘i are heavily invaded by non-native tree species. Such species constitute 86% or more of large trees, saplings, and seedlings in mesophytic forests; 45% of large trees and 66% of seedlings in their montane rainforests.
The most abundant tree species in Hawai`i is the invasive species Psidium cattleyanum (strawberry guava). It was recorded on 88, or37%, of 238 FIA plots. There are nearly twice as many P. cattleyanum saplings as Hawai`i’s most widespread native species, ‘ohi’a lehua (Metrosideros polymorpha).
Widescale replacement of native trees by non-native species is likely. Several factors favor these changes: 1) tree disease – rapid ‘ohi’a death has had drastic impacts on ‘ohi’a populations on several islands; 2) invasions by forbs and grasses; 3) soil damage and other disturbances caused by invasive ungulates; and 4) climate change. If succession conforms to these trends, non-native tree species could eventually constitute 75% or more of the forest tree stems and basal area on all islands and across forest types and elevations.
Loss of Hawai’i’s native tree species would be disastrous for biodiversity at the global level. More than 95% of native Hawaiian tree species are endemic, occurring nowhere else in the world.
The authors analyzed plant presence data from 238 FIA plots. Plots spanned the state’s various climates, soils, elevations, gradients, ownership, and management. However, access issues precluded inclusion of forests from several islands: Moloka‘i, Kaho’olawe, and Ni‘ihau. I know that Moloka‘i, at least, has a protected forest reserve (a Nature Conservancy property) at the island’s highest elevations.
Protecting Native Trees
Federal, state, and private landowners have carried out numerous actions to protect native forests. These efforts might be having some success. For example, forests on public lands, in conservation reserves, or in areas fenced to exclude ungulates were less impacted by non-native plants than unfenced plots, on average. However, the authors could not determine how much of this difference was the result of management or because protections were established in forests with the lowest presence of IAS species. Fencing did not prevent invasions by forbs and grasses – possibly because they are so widespread that seed sources are everywhere.
Hawaii’s two National parks (Hawai`i Volcanoes and Haleakala) have made major efforts to control invasive plants. Hawai`i Volcanoes, on the Big Island, began its efforts in the 1980s; Haleakala (on Maui) more recently. This might be one explanation for the fact that a smaller proportion of the forests on these two islands have been invaded. These efforts have not fully protected the parks, however. Low elevation native rainforests now have a high presence of non-native shrubs. Such forests on Hawai`i Island also have significant invasions by non-native woody vines, forbs and grasses.
More discouraging, intensive efforts have not returned lowland wet forest stands to a native-dominated state. Native tree species are not regenerating—even where there is plentiful seed from native canopy trees and managers have repeatedly removed competing non-native understory plants.
Potter et al. conclude that other approaches will be needed. They suggest deliberate planting of native and non-invasive non-native species or creation of small artificial gaps that might facilitate recovery of native tree species. In montane forests on Hawai`i and Maui, where native tree seedlings account for more than 70% of all tree seedlings, they propose enhancing early detection/rapid response efforts targetting invasive forbs. This would include both National parks.Certainly Haleakala National Park has this priority in mind. It launched a serious effort to try to eradicate Miconia calvescens when this tree first was detected.
Lloyd Loope, much-mourned scientist with US Geological Survey, attacking Miconia on Maui
Potter et al. note the challenge of managing remnant xerophytic dry forests, where natural regeneration of native plants has been strongly limited by invasive grasses; loss of native pollinators and seed dispersers; and the increasing frequency and intensity of droughts. They note that expanded management efforts must be implemented for decades, or longer, to be successful.
Native Trees at Risk to Nonnative Insects
Beyond the scope of the Potter et al. study is the fact that at least two dry forest endemic trees have faced their own threats from non-native insects.
The Erythrina gall wasp, Quadrastichus erythrinae, appeared in Hawai`i in 2005; it originates in east Africa. It attacks the endemic tree, wiliwili, Erythrina sandwicensis. I believe a biocontrol agent, Eurytoma erythrinae, first released in 2008, has effectively protected the wiliwili tree, lessening this threat.
The Myoporum thrips, Klambothrips myopori, from Tasmania, was detected on the Big Island in 2009. It threatens a second native tree. Naio, (Myoporum sandwicense), grows in dry forests, lowlands, upland shrublands, and mesic and wet forest habitats from sea level to 3000 m. The loss of this species would be both a signifcant loss of native biodiversity and a structural loss to native forest habitats. The thrips continues to spread; a decade after the first detection, it was found on the leeward (dry) side of Hawai`i Island with rising levels of infestation and tree dieback.
Rhus sandwicensis on Maui; photo by Forrest and Kim Starr
Two native shrubs, Hawaiian sumac Rhus sandwicensis and Dodonea viscosa, might be at risk from a biocontrol agent in the future. APHIS has approved a biocontrol for the highly invasive Brazilian pepper, Schinus terebinthifolia. Brazilian pepper is the second-most abundant non-native tree species in the State. It was found on 28 of 238 (12%) FIA plots. However, the APHIS-approved biocontrol agent is a thrips—Pseudophilothrips ichini. It is known to attack both of these two native Hawaiian shrubs. The APHIS approval allowed release of the thrips only on the mainland US. However, many insects have been introduced unintentionally from the mainland to Hawai`i. Furthermore, Hawaiian authorities were reported to be considering deliberate introduction of P. ichini to control peppertree on the Islands.
In Conclusion
In conclusion, Potter et al. found that most Hawaiian forests are now hybrid communities of native and non-native species; indeed, a large fraction are novel forests dominated by non-native trees. Business-as-usual management will probably mean that the hybrid forests – and probably those in which the canopy is currently dominated by native species—will follow successional trajectories to novel, non-native- dominated woodlands. This likelihood results in a more dire future for native plants in Hawaiian forests than has been previously described.
Potter at al. hope that their findings can guide research and conservation on other islands, especially those in the Pacific. However, Pacific islands already have the most naturalized species globally for their size—despite what was originally considered their protective geographic isolation.
SOURCE
Potter, K.M., C. Giardina, R.F. Hughes, S. Cordell, O. Kuegler, A. Koch, E. Yuen. 2023. How invaded are Hawaiian forests? Non-native understory tree dominance signals potential canopy replacement. Landsc Ecol https://doi.org/10.1007/s10980-023-01662-6
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Ponderosa pine, Coconico National Forest; photograph by Brady Smith, USFS
Have you noticed, as I have, a spurt of interest in conservation of trees? I can rejoice that more people now focus on this!!!!
I have blogged previously about international and national efforts to determine not only native species deserving conservation priority – by the Morton Arboretum and IUCN but also species most threatened by non-native pests. I have also reported on growing attention to breeding tree resistance to non-native pests.
Some scientists are now focusing on species’ regeneration as a way to understand the probable future of both native and introduced species. I hope that scientists will integrate these new data with existing information on the impacts of invasive non-tree plants and tree-killing introduced pests. We need such a comprehensive picture. That will be a challenge!
Also, I hope attempts to set conservation priorities will influence decisions by governmental and non-governmental funders – and those who influence them! So far, I see little evidence that these key players are paying attention. Some Forest Service scientists and academics are pushing for expanded resistance-breeding efforts. Others are writing sophisticated analyses of non-native pests’ ecosystem impacts. But is the USDA leadership supporting stronger pest-prevention measures? Or funding for research on restoration of species? Are conservation NGOs addressing introduced forest pests?
Here, I summarize new work by Kevin Potter and his colleagues, published in two papers (full references at the end of this blog). After reading my summary, I’d like to know: What do you think? Do you agree with the focus on individual species’ regeneration to set conservation and control priorities? Do you agree with the priority species and geographic regions they suggest?? How should we resolve inconsistencies compared to the priorities suggested by the IUCN and Morton Arboretum? If you do agree, how would you suggest we move forward? If not, what approach do you think would be more useful?
A New Approach to Evaluating Species at Risk
Potter and Riitters (2022) point out that a species’ successful regeneration is key to its population’s future genetic diversity. That, in turn, determines the organisms’ ability to adapt to environmental stress and change. The latter includes, but is not limited to, climate change. Because trees are immobile and long-lived, their populations probably require substantially more genetic variation than those of other kinds of plants.
Potter and colleagues (both articles) used FIA survey data to examine regeneration rates by both tree species native to the continental United States (= CONUS) and non-native tree species introduced to CONUS, Hawai`i, or Puerto Rico. I rejoice that they have included these tropical islands, which are part of the United States and are centers of plant endemism. (Two other blogs provide details on their findings in Hawai`i and Puerto Rico.
Native Trees at Risk: Focus on Poor Regeneration
For CONUS, Potter and Riitters (2022) asked whether 280 native forest tree species are regenerating at sustainable levels, both across their full ranges and in regional portions of their ranges, defined by provisional seed zones (an area within which plant materials are assumed to be adapted). Tree species for which FIA surveys placed 75% of the stems in the sapling or seedling classes are determined to be regenerating at sustainable levels. Tree species exhibiting lower proportions of their stems in these “small tree” classes are said to be failing to regenerate adequately.
Potter and Riitters (2022) found that 46 of the 280 native tree species (16.4%) might be at risk of losing important levels of genetic variation (see the list of species in Table 2 of the article). These included high proportions of species evaluated in the following genera: two of three Platanus species; two of four Nyssa species; about 40% of Juniperus and Pinus; and five of 46 Quercus species (10.9%).
[Many areas of the eastern forest, especially in the Mid-Atlantic region, are reported by Stout, Hille, and Royo (2023) to be have insufficient advance regeneration to replace canopy trees.]
Some species appear to be headed toward outright extinction, not only loss of genetic diversity. These include four relatively rare species in California: Pinus muricata, Platanus racemosa, Pseudotsuga macrocarpa, and Sequioadendron giganteum.No seedlings or saplings are recorded on the plots on which they occurred. I note that Platanus racemosa in southern California is being attacked and killed by the Fusarium dieback vectored by the polygamous and Kuroshio shot hole borers.
Platanus racemosa riddled by invasive shot hole borer; photo by Beatriz Nobua-Behrmann, University of California Cooperative Extension
I find it alarming that a few of the possibly at-risk species have extremely wide distributions. These are Populus deltoides (eastern cottonwood), Platanus occidentalis (American sycamore), and ponderosa pine (Pinus ponderosa). Another group of species are classified as at potential risk in all their seed zones: Juniperus californica, Juniperus osteosperma, Pinus pungens, and Quercus lobata (valley oak). I note that valley oak is also under attack by the recently introduced Mediterranean oak borer. Its vulnerability is exacerbated by its relatively small range.
Potter and Riitters (2022) found distinct geographic hot spots: 15 at-risk species occur primarily in the Southeast and 14 species are in California; both represent nearly a third of the at-risk species.
In general, high rates of regeneration failure are seen in the West. Nine at-risk species (19.6% of the 46) grow in the Southwest, eight in Texas (17.4%), and four in the Rocky Mountains (8.7%). However, the Northeast and Midwest are not immune. Seven species from the former and six from the latter are also regenerating poorly. Considering pines alone, seven of 14 at-risk speciesare in the West and five in the Southeast.
Seed Zones: a Proxy for Local Genotypes
As I noted at the beginning, Potter and Riitters (2022) used USDA Forest Service provisional seed zones as a proxy for areas in which a species is presumably locally adapted. In addition to the 46 species considered failing to regenerate adequately throughout their entire ranges, Potter and Riitters (2022) determined that another 39 species are at potential risk of losing locally adapted genotypes. That is, their regeneration levels fell below the threshold in at least half of the seed zones in which they occurred. These potentially at-risk species are in the same taxonomic groups: 13 pines (33.3% of the 39 species in the category), six junipers (15.3%), and three oaks (7.7 %). These, too are concentrated in the Southeast and California: 40% are in the former — including both bald-cypress species — and 30.8% are in California. Another seven species (17.9% of the 39) are in Texas. The Midwest is home to seven species, the Northeast and Southwest each has five species (12.8%), and the Rocky Mountain region has three species (7.7%).
Bald-cypress; photo by Kej605 via WikiMedia
The seed zones with the largest numbers of species regenerating poorly are in the East, specifically the central Great Lakes region, western New York and Pennsylvania, along the Mid-Atlantic and New England coasts, and the coastal plain from southern South Carolina to eastern Texas. Potter and Riitters (2022) say these areas have such high numbers of at-risk species because they are home to so many tree species. I note [although Potter and Riitters (2022) do not] that these regions have also experienced severe levels of tree mortality due to the emerald ash borer (mature and young trees), beech leaf disease (primarily young trees), and laurel wilt disease (sub-canopy trees).
A different geographic pattern appears when considering the proportion — rather than the number — of species facing deficits in regeneration. In several Western regions, 60 – 100% of the tree species fell below the study’s threshold of 75% of recorded stems being in the sapling or seedling sizes. These seed zones are found particularly in parts of California, the Southwest, the Great Basin, and the Pacific Northwest. In none of the seed zones in the East are more than 50% of tree species in the category of potentially losing genetic variation. The implication is that while more species might be lost from parts of the East, the loss of fewer species in some Western seed zones could result in larger impacts on the composition, structure, and function of forest ecosystems there.
Potter and Riitters (2022) say that their approach has limitations because it relies on an assumption that a lack of smaller (i.e., younger) trees is an indication that a species has inadequate regeneration across all or part of its distribution and thus is vulnerable to losing genetic variation. They are not able to quantify directly the genetic variation within most forest tree species. In addition, the choice of 75% or fewer of all trees being seedlings or saplings threshold as the threshold is arbitrary. They believe these decisions are defensible.
Potter and Riitters (2022) hope that indicators of forest sustainability such as this can bridge the gap between scientists, forest managers, policy makers, and other stakeholders.
Further, the authors hope that this approach will help prioritize species most in need of: 1) monitoring for genetic diversity, 2) in situ conservation, and 3) ex situ propagule collections. In a future blog I will compare the species highlighted by Potter and Riitters (2022) to the earlier priority list developed by the IUCN and Morton Arboretum. Finally, the focus on regeneration levels could help scientists design representative sampling protocols for range-wide ex situ propagule collections for genetic diversity studies using molecular markers.
Applying This Analysis to Invasions by Non-native Trees
In a second study, Potter, Riitters, and Guo (full citation at end of this blog) flipped the focus: they used the same approach to quantify the degree of invasion by non-native trees in the U.S. I’ve blogged about this study, in general, here. Also see my separate blogs for its welcome application to Hawai`i and Puerto Rico.
Again, Potter, Riitters, and Guo hope their approach will assist in the crucial, difficult task of distinguishing between high-impact and less threatening non-native species. They warn, however, that the FIA survey procotol does not suit the needs of an early detection system.
Differentiating Invasive Tree Species’ Impacts
Potter, Riitters, and Guo note that thousands of non-native tree species have been planted around world to provide an extensive list of ecosystem services. Globally, 400 tree species have been recognized as naturalized (= consistently reproducing) or invasive (= spreading) in areas outside their native ranges. Contrary to some expectations, even relatively undisturbed forests are affected by invasive plants. In the continental United States, many fewer invasive plant species are trees than other forms/habits – shrubs, forbs, gramminoids. On the tropical islands, a much higher proportion of invasive plants are trees.
Lugo et al. (2022; full citation at end of this blog) find non-native tree species occupy a tiny fraction of the forest area of the continental United States [= CONUS], i.e., only 2.8% of the area, and only 0.4% of all tree species recorded in the FIA plots. However, these non-native tree species are widespread. They are found in 61% of forested ecosections in CONUS. Also, they are becoming more common in invaded sites. [Ecosections are divisions within 37 ecological provinces in the hierarchical framework developed by Cleland et al. (2007). There are 190 ecosections in U.S. forest biomes.]
Potter, Riitters, and Guo categorized those non-native tree species with at least 75% of stems detected by FIA surveys to be in sapling or seedling size as highly invasive. In other words, these species are successfully reproducing after reaching the canopy. So they might be more likely to alter forest functions and ecosystem services than those reproducing less robustly. They classified as species with 60 – 75% of recorded stems in these “small tree” categories as “moderately invasive.”
Potter, Riitters, and Guo suggest that control might more productively target the moderately invasive species in geographic regions where they have spread less so far – so presumably fewer seed-bearing mature specimens are present. They list as examples Picea abies, Pinus sylvestris, and Paulownia tomentosa.
In CONUS, FIA protocols specify reporting of 30 non-indigenous tree species.
Acer platanoides
Ailanthus altissima
Albizia julibrissin
Alnus glutinosa
Castanea mollissima
Casuarina lepidophloia
Cinnamomum camphora
Citrus sp.
Elaeagnus angustifolia
Eucalyptus globulus
Eucalyptus grandis
Ginko biloba
Melaleuca quinquenervia
Melia azedarach
Morus alba
Paulownia tomentosa
Picea abies
Pinus nigra
Pinus sylvestris
Populus alba
Prunus avium
Prunus persica
Salix alba
Salix sepulcralis
Sorbus aucuparia
Tamarix spp
Triadica sebifera
Ulmus pumila
Vernicia fordii
About half of these –16 species – qualified under the Potter, Riitters, and Guo criteria as highly invasive: Acer platanoides, Ailanthus altissima,Albizia julibrissin, Cinnamomum camphora, Elaegnus angustifolia, Melia azedarach, Melaleuca quinquenervia, Morus alba, Picea abies, Pinus nigra, Prunus avium, Salix alba, Salix sepulcralis, Triadica sebifera, Ulmus pumila, Vernicia fordii. An additional four taxa are ranked as potentially highly invasive: Tamarix; Eucalyptus grandis and E. globulus, Populus alba.
ring-billed gulls eating berries of Chinese tallowtree (Triadica sebifera); photo by TexasEagle via Flickr
I ask : Do YOU agree that these taxa are the most important to be tracking as potentially invasive in forests of the continental United States?
Potter, Riitters, and Guo distinguish between the most “common” and the most “widespread” invasive tree species – although they do not define the differences. Some of the most “common” or “widespread” species are not a surprise: Ailanthus altissima, Triadica sebifera (syn. Sapium sebiferum), and Acer platanoides. Ailanthus is categorized as highly invasive in 39 of 44 ecoregions in which it occurs. It is also notoriously difficult to manage. Triadica sebifera is classified as highly invasive in every one of the 20 ecoregions in which it occurs. It produces prolific seed crops that are widely dispersed by birds and water. It can invade both disturbed and undisturbed habitats. Some of the common or widespread species do surprise me: Ulmus pumila, Morus alba and Picea abies.
Most of the non-native tree species occur on only 2% of plots in the ecoregions in which they occur. However, some highly invasive trees exceed this level:
Triadica sebifera is detected on 8.6% of plots on average across 20 ecoregions;
Ulmus pumila is detected on 3.7% of plots across 39 ecoregions;
Elaeagnus angustifolia is detected on 3.3% of plots in 13 ecoregions;
Melaleuca quinquenervia is detected on 2.7% of plots in 4 ecoregions.
A. altissima is detected on only 2% of plots in the 44 ecoregions. This is surprising to me. I see it everywhere in the Mid-Atlantic – and elsewhere!
[In USFS Region 9 (24 states in the Northeast and Midwest), FIA surveys in 2019 detected Ailanthus on only 3% of plots, Norway maple and Siberian elm each on only 1% of plots (Kurz 2023).]
Eastern U.S. forests are invaded at rates several times those in Western forests, both as a proportion of plots that are invaded and the diversity of plant growth forms. The probability of invasion is highest in Eastern forests that are relatively productive and located in fragmented landscapes that contain developed or agricultural land. Non-native invasive trees are most prevalent along the Gulf Coast and in Mid-Atlantic and Midwestern States. Highly invasive non-native trees are most diverse in the ecoregions of the Mid-Atlantic and Southeast. I note that these regions also rank high in numbers of native tree species determined by Potter et al.’s other study to be reproducing an unsustainable levels.
The study found that non-native trees are almost entirely absent from the Rocky Mountain States and Alaska. However, I have seen Ailanthus in riparian areas of Utah, Arizona, and New Mexico. While few non-native tree species are recorded from ecoregions along the Pacific Coast, those areas are heavily invaded by other types of plants. Lugo et al. say those shrubs and forbs are not interfering with forest regeneration. Do YOU agree?
BLM & USFS botanists removing Spanish broom from Rogue River Canyon; photo by Stacy Johnson, BLM
On tropical islands included in the study – Hawai`i and Puerto Rico – the situation is very different. Together, these islands’ tree canopy covers less than 0.5% that of the area in the lower 48. Hawai`i is recognized as a global hotspot of non-native species richness. Naturalized non-native plant taxa constitute about half of the Hawaiian flora. The US Forest Service tracks twice as many non-native tree species in Hawai`i (62) than over the entire continental U.S. plus Alaska.
Of these 62 species, Potter, Riitters, and Guo identified 26 tree species as either highly or moderately invasive, either already or potentially highly invasive, three as moderately invasive, seven as potentially moderately invasive. In general, the richness of non-native tree species is higher in lower-elevation ecoregions, especially the lowland/leeward dry and mesic forests on O’ahu and lowland wet and mesic forests of the Big Island. [The article makes a brief reference to the probable role of rapid ʻōhiʻa death opening the canopy of the mesic and wet forests, thereby facilitating plant invasions.]Most Hawaiian ecoregions, especially those on O’ahu and Hawai’i Island, had higher non-native tree species richness than even the most highly invaded ecoregions in the lower 48 states. Parts of O’ahu & Maui had the most non-native tree species classified as highly invasive.
The Caribbean archipelago – including but not limited to Puerto Rico – has a lower proportion of non-native plant species than Hawai’i — 17% of plant species are not native. However, their presence is even higher: two-thirds of Puerto Rico’s forests comprise novel tree assemblages. This is probably because Puerto Rico has half the land area of the Hawaiian archipelago and has been part of global trade networks for 500 years instead of 200. Potter and colleagues identified 17 non-native tree species as highly invasive, 16 as potentially highly invasive, and two as moderately invasive.
On the continent only seven of 30 non-native tree species occurr on at least 2% of FIA plots across the ecoregions in which they are inventoried. Hawai’i is stunningly different: 56 of 62 species occurr on at least 2% of plots across ecoregions on average; 24 species are present on at least 10% of plots on average. One species, Psidium cattleyanum, is present on nearly half of surveyed plots across 13 ecoregions! In Puerto Rico, 21 species occurred on at least 2% of the FIA plots.
Acacia confusa – highly invasive in dry forests of Hawai`i; photo by Forrest and Kim Starr
Potter, Riitters, and Guo could not assess the invasiveness of several species that occurred only as small stems in a couple of plots. There are 11 such species on Hawai`i, eight on Puerto Rico. These species might be in the early stages of widespread invasion, or they might never be able to reproduce and spread. Despite the uncertainty, the authors suggest that eradication or control efforts targetting these species might be more cost-effective since presumably few trees have reached reproductive age yet. In Puerto Rico, they single out Schinus terebinthifolius, since it is already recognized as moderately invasive in Hawai`i [I add – seriously invasive in nearby Florida!]. However, they also emphasize the threat from one of the widespread species, Syzgium jambos, because it is a shade-tolerant species that can form dense, monotypic stands under closed canopies
I have posted separate blogs providing more details on the invasive tree species in Hawai`i and Puerto Rico.
Limits of the FIA Dataset
As in the study of native species regeneration, Potter, Riitters, and Guo specify limits arising from use of the FIA dataset. Two seem particularly pertinent to evaluation of the situation on the tropical islands.
First, they cataloged only those non-native tree species chosen by the FIA program administrators to track in the three major regions. Again, I ask YOU whether you agree with the species being recorded. Should others species be included? Should some of these species be dropped?
Second, the survey protocol does not differentiate between sites with significantly different status and history. For example, non-native trees growing on abandoned agricultural sites are counted the same way as those growing in presumably old-growth forests. They conclude that including such sites might explain the records of Eucalyptus and pine species in surveys on the islands.
Finally, as noted in the other study, the program incorporates plots that contain at least 10% canopy cover by live trees or had such cover in the past. The inventory has not included urban parks – although in recent years an urban inventory protocol has been developed.
I remind you that Potter, Riitters, and Guo warned that the FIA inventory is not designed to detect newly introduced species that are early in the invasion process.
SOURCES
Kurtz, C.M. 2023. An assessment of invasive plant species in northern U.S. forests. Res. Note NRS-311. http://doi.org/10.2737/NRS-RN-311
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, and C.M. Kurtz. 2022. The Contribution of NIS Tree Species to the Structure and Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific & Caribbean. USDA USFS General Technical Report IITF-54.
Potter, K.M and Riitters, K. 2022. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests 2022, 13, 19. https://doi.org/10.3390/f13010019.
Potter K.M., Riitters, K.H. and Guo, Q. 2022. Non-native tree regeneration indicates regional and national risks from current invasions. Frontiers in Forests and Global Change doi: 10.3389/ffgc.2022.966407
Stout, S.L., A.T. Hille, and A.A. Royo. 2023. Science-Management Collaboration is Essential to Address Current & Future Forestry Challenges. IN United States Department of Agriculture. Forest Service. 2023. Proceedings of the First Biennial Northern Hardwood Conference 2021: Bridging Science and Management for the Future. Northern Research Station General Technical Report NRS-P-211 May 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Asian longhorned beetle in South Carolina; photo by R. Brad Thompson, USDA APHIS
In March I asked your help in asking Congress to fund USDA programs that protect forests from non-native insects and pathogens. The Congress has now taken major steps to specify funding for Fiscal Year 2024 – which begins on 1 October. Both the House and Senate Appropriations committees have adopted their bills. They differ substantially. When Congress returns from its August recess in September, it will face many difficulties in negotiating the final spending levels – not just the different funding levels but also attached “riders” dealing with social and political issues, most of which have nothing to do with invasive species. [Some of the riders to seek to restrict application of the Endangered Species Act to several species, e.g., sage grouse and grey wolf.]
USDA APHIS
As you know, USDA’s Animal and Plant Health Inspection Service (APHIS) is responsible for preventing introduction of pests that harm agriculture, including forests; and for immediate efforts to eradicate or contain those pests that do enter. While most port inspections are carried out by the Department of Homeland Security Bureau of Customs and Border Protection, APHIS sets the policy guidance. APHIS also inspects imports of living plants. In the table below, I provide information on funding for key APHIS programs in FY23, the Administration’s request for FY24, the funding level the Center for Invasive Species thought necessary, and the House and Senate funding levels.
The earlier blog link has additional information: the FY22 funding levels and a fairly long justification for funding these APHIS programs. I never posted a blog discussing USFS funding due to my trip to Europe.
Appropriations for APHIS programs (in $ millions)
Program
FY 2023
FY 2024 Pres.’ request
CISP ask
House bill
Senate bill
Tree & Wood Pest
$63
$64
$65
$55.6
$62.6
Specialty Crops
$216
$222
$222
$224.5
$217.9
Pest Detection
$29
$30
$30
?
?
Methods Development
$23
$23
$25
?
$21.8
Emerg. Preparedness
$44
$45.2
NA
$44.6
$48
Contingency fund
$514
$543
$514
$514
I have not seen a report from the House Committee so I don’t know whether that body prioritized any invasive species issues.
The Senate report included this statement re: Sudden oak death
“The European strain 1 [EU1] and the North American strain 1 [NA1] of the sudden oak death pathogen are major threats to western Douglas-fir/tanoak forests, resulting in quarantine restrictions that threaten U.S. forests and export markets for log shipments and lily bulbs. The Committee recommendation includes no less than the fiscal year 2023 funding level to improve understanding of EU1 and NA1 strains of the sudden oak death pathogen and treatment methods to inform control and management techniques in wildlands.”
Appropriations for USFS programs (in $ millions)
Program
FY 2023
FY 2024 Pres.’ request
CISP ask
House bill
Senate bill
Forest Health Protection
Federal Lands
$32
$17
Coop Lands
$51
$33
Research & Development
$349.1
$349.1
$275
$307.3
Forest Inventory
$30.2
$30.2
$32.2
$32.2
Work on 10 invasive spp
$.5
$4.4
$8.5*
0
0
* CISP ask was intended as first step to increasing funding for invasive species to 5% of R&D funds. See the March blog for an explanation.
SOD-killed tanoaks in southern Oregon; Oregon Department of Agriculture photo
Senate report =Sudden Oak Death. “Since 2001, USFS has been treating SOD infestations on public lands in Oregon and California, in cooperation with Bureau of Land Managemebt. The Committee expects USFS continue these efforts; it provides $3M for SOD treatments and partnerships with States and private landowners.”
Urban & Community Forestry.—”The bill provides $40 M for urban and community forestry. The Committee recognizes the critical need to restore and improve urban forests due to dominance of exotic invasive woody species. USFS should prioritize regional multi-organizational collaborations to support conservation efforts that help trees adapt to and offset climate change, which model best practices for effective urban and community forestry grants. The Committee also expects the program to prioritize tree-planting in socially disadvantaged and historically underserved communities with low canopy coverage, including Tribal communities.”
Summary of Justifications: The Costs of Introduced Pests
Introduced pests threaten many forest products and ecosystem services benefitting all Americans. Already, the 15 most damaging non-native pests threaten at least 41% of forest biomass in the “lower 48” states. In total, these 15 species have caused an additional annual conversion of live biomass to dead wood at a rate similar in magnitude to that attributed to fire (5.53 TgC per year for pests versus 5.4 to 14.2 TgC per year for fire). Fei et al.; full citation at end of blog.
These pests also impose significant costs that are borne principally by municipal governments and homeowners. As more pests have been accidentally introduced over time, these costs have risen.
Pathways of Introduction
The many tree-killing wood-boring pests arrive in inadequately treated crates, pallets, and other forms of packaging made of wood. The March blog presents 2023 data on import volumes and the findings of Haack et al. 2022. The point is, ISPM#15 has fallen short. [See blogs under “wood packaging” category on this site]
APHIS’ Tree and Wood Pests account supports eradication and control efforts targeting principally the Asian longhorned beetle and spongy (= gypsy) moth. Eradicating the ALB normally receives about two-thirds of the funds. The programs in Massachusetts, New York, Ohio, and South Carolina must continue until eradication succeeds. The emerald ash borer continues to spread since APHIS dropped regulations attempting to halt this. EAB was detected in Oregon in 2022; and on Colorado’s western slope in 2023.
Other pests—especially plant diseases like sudden oak death and sap sucking insects like hemlock woolly adelgid—come on imported plants. I noted that no studies have examined the risk of pests arriving on the ~5 billion plants we Americans now import annually (see March blog). The information gap is particularly alarming regarding pathogens. Evidence of failures:
the recent detection of two strains of Phytopthora ramorum that were formerly limited to Europe in forests of Oregon and California;
and the apparent introduction of the causal agent(s) of beech leaf disease.
APHIS manages damaging pests introduced on imported plants or other items through its Specialty Crops program. The principal example is its efforts to prevent spread of the SOD pathogen through the interstate trade in nursery plants. I am pleased that the Senate report calls on APHIS to focus on that pathogen’s growing genetic diversity.
Beech leaf disease has spread >700 miles since its first detection just 11 years ago
The Administration did not persuade the Congress to fund a $1 million emergency fund for APHIS – although they did fund both “emergency” and “contingency” programs. I am not certain about the difference.
Furthermore, both chambers of Congress included in their legislation – not in the report – language instructing the Secretary of Agriculture to use his authority to obtain emergency funds from other USDA agencies to address animal or plant health emergencies:
“Provided further, That in addition, in emergencies which threaten any segment of the agricultural production industry of the United States, the Secretary may transfer from other appropriations or funds available to the agencies or corporations of the Department such sums as may be deemed necessary, to be available only in such emergencies for the arrest and eradication of contagious or infectious disease or pests of animals, poultry, or plants, and for expenses in accordance with sections 10411 and 10417 of the Animal Health Protection Act (7 U.S.C. 8310 and 8316) and sections 431 and 442 of the Plant Protection 15 Act (7 U.S.C. 7751 and 7772), and any unexpended bal1ances of funds transferred for such emergency purposes in the preceding fiscal year shall be merged with such transferred amounts.”
The Congress has included this or similar language in appropriations reports for almost two decades, but it has not succeeded in freeing up many funds for countering plant pests. Perhaps placing the language in the legislation rather than the report will help … we will have to see.
In the meantime, I have been working with others to amend the Plant Protection Act to ensure that the emergencies so referenced include threats to forests. See §2 in H.R. 3174 link (Balint, Vermont) and S. 1238 (Welch, Vermont).
SOURCES CITED
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice .2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
infested wood bearing ISPM#15 mark; photo by Oregon Department of Agriculture
SWPM has been recognized as a major pathway for introduction of tree-killing pests since the Asian longhorned beetle was detected in New York and Chicago in late 1990s. As of 2014, 58 new species of non-native wood- or bark-boring insects had been detected – many probably introduced via wood packaging [Leung et al. 2014]. Other examples include the emerald ash borer, redbay ambrosia beetle, and, possibly, the invasive shot hole borers.
In response to recognition of the pest risk associated with wood packaging, countries adopted ISPM#15. This process was reviewed in the two articles by Haack et al. and my recent blog. I provided the broader context of the World Trade Organization (WTO) in my Fading Forests II report.
I have blogged often about the continuing poor compliance with wood packaging regulations, especially by China; and USDA APHIS’ insufficient efforts to fix the problems. The DHS Bureau of Customs and Border Protection (CBP) has tried much harder. See particularly my blog about Bob Haack’s re-evaluation of the pest approach risk in wood packaging. Given the high volumes of imports, pests infesting even a small proportion of incoming shipments can result in tens of thousands of pest-infested containers entering the U.S. or Canada each year. For an explanation of these calculations, see the “background” section of this blog.
Since 2010, CBP has discovered actionable pests in more than 700 shipments each year (pers. comm.). [APHIS reports half as many detections – 300 wood boring and bark beetles (Greenwood et al. citing APHIS report from 2021). Perhaps the difference arises from some of the actionable pests not being wood-borers, e.g., snails.] The persistence of pest presence has disappointed CBP staffers, because the agency has taken several actions intended to discourage violations. These include imposing fines and revoking the violators’ participation in the U.S. Trade Partnership Against Terrorism (C-TPAT) program. Greenwood et al. describe these consequences of non-compliance, as well as the expense of re-exporting the goods and associated wood packaging, as “significant”. Regardless of how significant they might be, so far these consequences have not reduced non-compliances substantially.
The fact is, countries cannot rely on the presence of the ISPM#15 mark or stamp to indicate that the wood packaging is pest-free. In both the United States and Europe, more than 90% of the SWPM found to be infested has born the ISPM#15 stamp (pers. comm.; Eyre et al. 2018). All the pest-infected shipments imported after 2006 discussed in the Haack et al. 2022 study were in wood packaging bearing the ISPM#15 mark. While many of the problems arise on shipments from Asia, findings occur sporadically with countries all across the globe- and notably, U.S. importers have also found serious problems with dunnage from Europe.
But that is the purpose of the standard!
Two outstanding questions that need answers
Continuing poor compliance with regulations by China. This is despite the fact that the U.S. and Canada have required treatment of wood packaging from China since December 1998 – nearly 24 years. Haack et al. found that the proportion of Chinese consignments with infested wood is five times greater than expected based on their proportion of the dataset. The rate of wood packaging from China that is infested has remained relatively steady: the Chinese infestation rate was 1.26% during 2003–2004, and ranged from 0.58 to 1.11% during the next three periods.
Why are the responsible agencies in the United States not taking more aggressive action to correct this long-standing problem? This is a matter of political will.
Despite the ISPM#15 mark being unreliable for more than a decade, countries have not carried out research to determine the root causes. Even now (i.e., Haack et al. 2022; Greenwood et al.) no one can say what proportion of these ISPM-marked but pest-infested pieces of wood results from the treatment not being effective in killing all pests; what proportion results from inadequate application of treatments that are per se effective; and what proportion from fraud (deliberate claims to have applied a treatment that was not done)?
Admittedly, answering these questions will not be easy. First, there is no independent test for whether treatments have been applied; the treatments do not alter the wood’s properties in measurable ways. Scientists need experiments to test the real-world efficacy of treatments in the specific contexts of solid wood packaging.
Second, each country is responsible for its own compliance. Countries differ in their capacity and political will to address this issue. However, success of ISPM#15 depends on determining the cause of continuing pest presence in wood marked as treated, and taking appropriate action to solve the underlying problem.
Greenwood et al. attempt to make progress toward carrying out this necessary task by describing the many steps in the wood packaging supply chain, associated opportunities for pests to infest the wood at each step, and actions exporters and importers can take to try to minimize the risk.
Again, as I discussed in the earlier blog, Haack et al. (2022) found several disturbing situations:
While the pest approach rate has fallen since U.S. implementation of ISPM#15, the extent of the decline has progressively decreased as time passes. The reduction during 2005–2006 was 61%; during 2007–2009, 47%; during 2010-2020 only 36%.
The 2010 – 2020 pest approach rate was calculated at 0.22%. This is more than double the rate based on 2009 data (0.1%, as stated in Haack et al. 2014). While we cannot directly compare these two data points (the two studies used different methods, as discussed in the blog), the bottom line is that the approach rate remains too high. Our forests continue to be exposed to the risk of introduction of highly damaging wood-boring pests. Furthermore, since the number of countries sending us infested wood packaging has increased, those potential pests include insects from a greater variety of countries (biomes).
The two most commonly intercepted families of wood borers are Cerambycidae and Scolytinae (Haack et al. 2022). These families include the Asian longhorned beetle, , redbay ambrosia beetle, and invasive shot hole borers. The 2009 amendment requiring debarking has not apparently resulted in substantial decreases in pest presence, although the proportion of pests that are true bark beetles has declined – from 100% of Scolytinae identified to genus or species detected before 2009 to only 23% in 2010–2020 period.
Michigan’s champion green ash killed by emerald ash borer
Haack et al. (2022) Recommendations
Haack et al. (2022) call for several improvements. Several pertain to how data are collected. Recording the number of infested pieces of wood instead of reporting only consignments would help clarify whether the numbers of insects reaching our borders has fallen, risen, or remained steady. Recording the presence of bark – and the size of any bark remnants – would help clarify whether pests are re-infesting treated wood.
They also note opportunities to improve ISPM#15 implementation and enforcement through training. However, compliance issues persist despite past educational efforts by APHIS and the IPPC.
The Wood Packaging Supply Chain Offers many Opportunities for Pests to Infest the Wood
Greenwood et al. describe each step in fabricating wood packaging material and the opportunities each step presents for unwanted organisms to enter that supply chain. They note that ensuring that these organisms are not then transported on wood packaging being used to carry goods requires that the pests be removed; rendered infertile, inactive, unable to complete development or reproduce; or killed.
The first step in fabricating wood packaging is to harvest trees. Those trees probably harbor various insects, fungi, nematodes, and other organisms that use trees as a resource — for food, shelter, or as a substrate for oviposition. Greenwood et al. mention that the multiplicity of organisms’ life histories pose different challenges for detection and management depending on size, type of tissue utilized, and other factors. The likelihood that a pest or pathogen will be present on or in tree tissues depends on several biotic and abiotic factors, including a species’ proclivity to experience periodic or episodic outbreaks; blow-down events (e.g., hurricanes, windstorms); and harvesting practices. Some of these factors can be controlled by people harvesting the wood.
One of the most frequent opportunities for pest infestation, escape, or cross-contamination is when the wood is stored in the environment. Such storage events happen after the tree is felled — at either the harvest site or processing facility; after the pallet or crate is built – either empty or after the goods have been packed; at the port of export before embarkation; at the importing port before inspection or onward transport; at distribution centers; at retailers; at “pallet graveyards” while awaiting repair or recycling. Retailers and customers have few resources for responsible handling of SWPM – and few incentives to be careful.
a “pallet graveyard”; photo by Adnan Prasad, then with Davey Tree
The risk is exacerbated if storage takes place near woodlands. photo from Savannah At ports and distribution centers, the presence of SWPM from many origins adds to the risk of cross-contamination. Enclosing the SWPM in containers does not completely eliminate the risk since organisms might enter through cracks or air vents. Greenwood et al. suggest management tactics to prevent or reduce pest interaction with the wood during these periods.
container storage near a treed area – Port of Savannah; photo by F.T. Campbell
One of the ISPM#15 requirements intended to minimize the pest risk is debarking the wood. This process removes most organisms that live in and just under the bark. However, debarked wood usually retains some patches of bark because trees are not perfectly round cylinders. Therefore ISPM#15 specifies that remaining bark must be less than 3 cm wide or, if the piece is longer than 3 cm, less than 50 cm2 in area.
Greenwood et al. state that after debarking and treatments per ISPM#15, the risk that a pest will be present on the SWPM has been significantly reduced. However, other challenges appear as the newly-minted packaging is put into use – primarily through the possibility of contamination during storage – as described above. There are also risks associated with inadequate or insufficient treatment or fraud.
Once loaded onto a ship, containers and any SWPM, including dunnage, are very difficult to inspect. That means that the loading process presents that last opportunity for inspection and mitigation of contaminating pests. Greenwood et al. note that it is the shipper’s responsibility to ensure containers are “clean, free of cargo residues, noxious materials, plants, plant products and visible pests” before being loaded on the ship. However, the International Maritime Organization (IMO) provides only recommendations, not mandates. Australia has adopted more stringent requirements.
Arrival at the importing country’s port presents the first opportunity for non-indigenous organisms to escape and the first domestic opportunity for the receiving country to inspect the shipment. While U.S. and Canadian customs agencies have authority to board ships before they dock to inspect them, Mexican agencies do not. The most extensive pre-docking requirements are aimed at preventing arrival of moths in the Lymantria genus from Asia.
dunnage in Houston; photo by S. Useman, CBP
Greenwood et al. note that dunnage presents unique risks. After it is removed from ships during the unloading process it is often stored at the port. As noted above, storage in the open allows pests to escape to nearby trees or to cross-contaminate other SWPM. Ports struggle to manage these piles. In 2016 the U.S. revised its regulations to allow for the more rapid destruction of illegally deposited dunnage via incineration at the port. Since 2008 Canada has considered all shipborne dunnage to be non-compliant – regardless of whether it bears the ISPM#15 stamp. In the largest Mexican ports, dunnage is fumigated and destroyed. However, the dunnage might be stored in the open for considerable periods before being destroyed.
Worse, it is often impossible to assign chain of custody information and responsibility for either disposition of non-compliant dunnage or penalties for non-compliance. Dunnage or blocking pieces might be added immediately before shipping by entities other than the owners or brokers for the commodities being shipped. I have already noted that it is nearly impossible to inspect dunnage in a ship’s hold.
Unfortunately, studies have not clarified the level of infestation of dunnage in comparison to other wood packaging types made from multiple pieces of milled wood, such as pallets or spools.
Greenwood et al. describe the different fates of pallets, dunnage, crates, spools, and other types of SWPM. Wood pallets are frequently recycled or remanufactured in the U.S., although there are no data on the proportion of the recovery market that is composed of pallets initially manufactured overseas. In the U.S., most repairs are done with components from reclaimed pallets so they probably conform to ISPM#15 repair guidelines. However, contamination could happen while the pallets are in storage awaiting reuse. As SWPM ages, different types of pests might be attracted.
SWPM deemed not suitable for reuse is either destroyed in controlled settings (i.e., solid waste facilities, wood processing facilities, or landfills), used in recycling or downcycling markets, or reclaimed. It might be chipped and sold as mulch, soil amendment, or animal bedding; or it might enter the commercial fiber market and be manufactured into other wood products (e.g., paper, chipboard, fuel pellets). These dispositions present very low pest risk, due to the final dimensions of the wood products being too small to sustain pest development in most cases. However, some microorganisms and very minute arthropods might persist even on chipped or shredded material. There is little data on the final disposition of SWPM globally.
Greenwood et al. reiterate that the presence of hitchhiking or contaminating pests does not imply failure of ISPM#15 treatments, which do not target such organisms. Such pests can also be present on non-wood packaging material such as plastic and metal. Countries vary in their concern about these hitchhiking pests, which include dry wood borers and brown marmorated stinkbug (Halyomorpha halys). Since these pests are not addressed by ISPM#15, countries can implement their own management strategies to counter contaminating pests on all SWPM, containers, and conveyances. Indeed, Pennsylvania regulates the movement of SWPM and other high risk articles to prevent the spread of the non-specific hitchhiking pest, spotted lanternfly, Lycorma delicatula.
They also note that reuse, disposal, and recycling of packaging made from metal, plastic, or even paper requires very different processes and facilities than those used for wood.
Greenwood Recommendations
Greenwood et al. advocate additional research on several questions:
to test whether currently accepted ISPM#15 treatments are sufficiently effective within the newly proposed metrics found in Ormsby 2022.
to determine the risk profile and enforcement of dunnage, especially whether organisms in dunnage are more likely to survive treatment (dunnage pieces are often much larger than any component piece of a pallet or crate).
to develop new treatments – including to counter re-infestation later in the supply chain. Scientists will probably have to replace Probit9 as a standard because it is not practical to exposing tens of thousands of wood-infesting insects to the new treatment. This is also discussed in Ormsby 2022.
to develop ways to test whether treatments have been applied – needed to verify whether fraud has occurred.
social and economic motivations around compliance
Most of these studies will require international cooperation.
Other steps are also need. As U.S. importers of break-bulk cargo have found out, procuring apparently compliant SWPM does not protect them from legal, financial, and logistical consequences if that SWPM turns out to be non-compliant or otherwise infested with live actionable pests. Some importers have begun exploring options toward additional private inspection at the exporting port, beyond solely requiring the use of ISPM#15 compliant materials. Greenwood et al. suggest the possibility of third-party certification. They also supported calls for officials to release of information about which foreign facilities have a history of selling SWPM subsequently found to be non-compliant. This information would empower importers to procure pest-free SWPM – thus harnessing market incentives to improve compliance.
Managing all this + pest risks? Photo by Port Authority of Long Beach
Greenwood et al. say that reducingexternal contamination on conveyances – ships, airplanes, trucks, and trains – is challenging. It would require the cooperation of multiple entities who manage yards, equipment, and facilities. Improved management must make sense to people who have severe constraints on time, staffing, space, and safety protocols. Persuading them to act will probably depend on improved information (research) on the cost effectiveness of various strategies and real-world incidence of contamination in different storage scenarios (beyond Lymantria complex), plus development of new surveillance tools.
Greenwood et al. suggest that conducting a HACCP assessment of the supply chain could help identify how a systems approach might better mitigate pest risks of SWPM. They think systems approaches might be especially promising for reducing risks of contaminating organisms. NAPPO recently adopted a standard for designing and implementing systems approaches for wood commodities.
Finally, I remind you of my recommendations for immediate policy actions to hold foreign suppliers responsible for non-compliant wood packaging:
U.S. and Canada should refuse to accept wood packaging from foreign suppliers that have a record of repeated violations – whatever the apparent cause of the non-compliance. They should institute severe penalties to deter foreign suppliers from taking devious steps to escape being associated with their violation record.
I also support the suggestion (above) that phytosanitary agencies inform importers on which foreign treatment facilities have a record of poor compliance or suspected fraud – so the importers can avoid purchasing SWPM from them.
U.S. and Canada should encourage importers to switch to materials that won’t transport wood-borers. Cardboard and manufactured wood packaging (e.g. oriented strand board and compressed wood block) are wood fiber products that have near zero risk of wood-borer infestation. Plastic is also one such material. I note that Earth is drowning under discarded plastic.
APHIS and CFIA have the authority to take these action under the “emergency action” provision (Sec. 5.7) of the World Trade Organization’s Agreement on the Application of Sanitary and Phytosanitary Standards (WTO SPS Agreement). (For a discussion of the SPS Agreement, go to Fading Forests II, here.)
Longer-term Actions
APHIS and CFIA should exercise their right to set a higher “level of protection” to minimize introductions of pest that threaten our forests (described inter alia here.) They should prepare a risk assessment to justify adopting more restrictive regulations that would prohibit use of packaging made from solid wood – at least from the countries with records of high levels of non-compliance.
The studies needed to determine the cause of the continuing issue of the wood treatment mark’s unreliability, and appropriate actions to fix the problem, should be conducted with other countries. Appropriate entities would be the International Plant Protection Convention (IPPC) and International Forest Quarantine Research Group (IFQRG). However, if attempting such collaboration causes delays, APHIS and CFIA should begin unilaterally.
Meanwhile, what can we do?
Urge Congress to conduct oversight on APHIS’ failure to protect America’s natural resources from continuing introductions of nonnative insects and diseases. Note that the Mediterranean oak borer has apparently been introduced several times in recent years – despite ISPM#15.
Raise the issue with local, state, and federal candidates for office;
Urge Congress to include provisions of H.R. 3174 / S. 1238 in the 2023 Farm Bill;
Ask any associations of which you are a member to join in communicating these concerns to Congressional representatives and senators. These include:
if you work for a federal or state agency – raise to leadership; they can act directly or through National Plant Board, National Association of State Departments of Agriculture, National Association of State Foresters, National Governors Association, National Association of Counties …
scientific membership societies – e.g., Society of American Foresters, Entomological Society of America, Phytopathological Society;
individual conservation organizations, either with state chapters or at the national level;
woodland owners’ organizations, e.g., National Woodland Owners Association, National Alliance of Forest Owners, and their state chapters
urban tree advocates
International Forest Quarantine Research Group
Write letters to the editors of your local newspaper or TV news station.
SOURCES
Eyre, D., R. Macarthur, R.A. Haack, Y. Lu, and H. Krehan. 2018. Variation in Inspection Efficacy by Member States of Wood Packaging Material Entering the European Union. Journal of Economic Entomology, XX(X), 2018, 1–9 doi: 10.1093/jee/tox357
Greenwood, L.F., D.R. Coyle, M.E. Guerrero, G. Hernández, C.J. K. MacQuarrie, O. Trejo, M.K. Noseworthy. 2023. Exploring pest mitigation research and management associated with the global wood packaging supply chain: What and where are the weak links? Biol Invasions https://doi.org/10.1007/s10530-023-03058-8
Haack, R.A., K.O. Britton, E.G. Brockerhoff, J.F. Cavey, L.J. Garrett, et al. 2014. Effectiveness of the International Phytosanitary Standard ISPM No. 15 on Reducing Wood Borer Infestation Rates in Wood Packaging Material Entering the United States. PLoS ONE 9(5): e96611. doi:10.1371/journal.pone.0096611
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice. 2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Frontiers in Forests and Global Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ. 2014. doi:10.1890/130311
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
I have blogged many times about the risk of pest introductions on imports of live plants [= “plants for planting” in USDA’s terms]. Last October I reviewed 14-year old data indicating that nearly 70% of 455 damaging tree pests introduced to the continental U.S. had probably been introduced via plant imports. These included 95% of sap feeding and 89% of foliage feeding insects and about half of the pathogens. The approach rate of pests on imported plants was apparently 12% (Liebhold et al. 2012) — more than 100 times higher than the 0.1% approach rate found by Haack et al. (2014) for wood packaging.
First, those analyses focus almost exclusively on insects (MacLachlan et al. 2022 focused on a single insect order, the Hemiptera!), despite the many pathogens probably introduced by the plant trade in recent decades. Examples I cited included several Phytophthoras, rapid ohia death, beech leaf disease, and boxwood blight. There have been repeated detections of the Ralstonia solanacearum Race 3 biovar 2.
SOD- infected rhododendrons; photo by Jennifer Parke, Oregon State University
Second, most studies analyzing the pest risk associated with plant imports use port inspection data – which are not reliable indicators of the pest approach rate – as explained by Liebhold et al. 2012 and Haack et al. 2014 (as it pertains to wood packaging).
Third, many of the studies are based on data from a decade or longer in the past. This means the studies do not address whether APHIS’ recent changes in its approach – including adoption of NAPPRA – have resulted in reduced introductions.
A complication is that, since insects are difficult to detect, those associated with the high volumes of plants imported in recent years might not be detected for years or decades after their introduction.
I have called for APHIS to update the Liebhold et al. 2012 study to determine the approach rate for all types of organisms that threaten North American tree species. Any such study should include trees on Hawai`i, Guam, Puerto Rico, and other U.S possessions and territories. These islands are nearly always excluded from analyses of imported pests. I concede that there are probably scientific and data-management challenges but these islands are immensely important from a biodiversity point of view, and they are parts of the United States!
eastern hemlocks killed by hemlock woolly adelgid; Linville Gorge; photo by Steven Norman, USFS
MacLachlan et al. (2022) estimated that new establishments – of insects in the order Hemiptera – per unit of additional plant imports have shrunk substantially. They attribute this decline to a combination of increased imports and the presence of a growing number of insect species introduced in the past. They found that introductions to the Asian Palearctic and Neotropic regions have been reduced by depletion of species pools. Other factors are thought to explain the substantial decline in establishment likelihood for the other regions. However, lag times in detecting insect introductions complicate this assessment.
However, despite that significant decrease in risk per unit of imports, MacLachlan et al. (2022) found that the number of establishments has remained relatively constant over the past century because of substantial increases in overall import levels and diversification of the origins of imports across regions, which exposed the U.S. to new source species pools.
MacLachlan et al. (2022) suggested that APHIS should target biosecurity resources to the specific commodity-country pairs associated with a higher relative risk of introducing additional insect species.
Recent studies are taking a welcome new stance: looking at links between introductions of non-native plant and insect species. I first raised this approach a year ago. Studies by teams led by Doug Tallany and Sara Lalk [Lalk et al.; articles by Tallamy] agree that:
Non-native plants – some of which are invasive – are altering ecosystems across broad swaths of North America and the impacts are insufficiently understood.
The invasive plant problem will get worse because non-native species continue to be imported, planted … and to invade.
Plant-insect interactions are the foundation of food webs – they transfer energy captured by plants through photosynthesis to other trophic levels, plus play a major role as pollinators. Consequently, changes to a region’s flora will have repercussions throughout ecosystems.
Dr. Tallamy studies the response of herbivorous insects to non-native woody plants – not just invasive plants, but also non-native plants deliberately planted as crops or ornamentals, or in forestry. Introduced plants have completely transformed the composition of plant communities in both natural and human-dominated ecosystems world-wide. The impacts can be significant: Burghardt et al. found that 75% of North American lepidopteran species and 93% of specialist species were found exclusively on native plant species.
monarch butterfly on milkweed; photograph by Jim Hudgins, USFWS
Lalk and colleagues studied the relationships between individual species of invasive woody plants and the full range of arthropod feeding guilds – pollinators, herbivores, twig and stem borers, leaf litter and soil organisms. They decry the absence of data on the complex interactions between invasive woody plants and arthropod communities at a time when invasive shrubs and trees are so widespread and causing considerable ecological damage. (See the blog for their specific research recommendations.)
Nor is the impact of non-native plants on insect fauna limited to North America. Outhwaite et al. found that the combination of climate warming and intensive agriculture is associated with reductions of almost 50% in the abundance and 27% in the number of species within insect assemblages relative to levels in less-disturbed habitats with lower rates of historical climate warming. These patterns were particularly clear in the tropics (perhaps partially because of the longer history of intensive agriculture in temperate zones). They found that high availability of nearby natural habitat (that is, native plants) can mitigate these reductions — but only in low-intensity agricultural systems.
Recognizing that plant diversity drives global patterns of insect invasion, Liebhold et al. (2023) compared various factors associated with numbers of invasive insect species in 44 land areas.They determined that the numbers of established non-native insect species are primarily driven by diversity of plants – both native and non-indigenous. Other factors, e.g., land area, latitude, climate, and insularity, strongly affect plant diversity; thus they influence insect diversity as a secondary impact. When I blogged about this study, I noted that the article appeared more than four years earlier, but has apparently had little influence on either policy formulation governing plant introductions or pest risk analysis applied to insects or pathogens that might be introduced. I suggested that we need a separate analysis of whether fungi, oomycetes, nematodes, and other pathogens show the same association with plant diversity in the receiving environment.
Studies of plant-insect relationships continue to be published. I welcome this!
Bonnamour et al. (2023) builds on the earlier studies. They also found that the presence of non-native plant species was a better predictor of insect invasions than such more widely discussed socioeconomic variables as trade volumes generally or even trade in plant products. However, detection of the associated insect invasions occurs years after detection of the plant invasions. Indeed, numbers of established non-native insect species corresponded more closely to plant introduction volumes in 1900 than current or recent import volumes.
Bonnamour et al. note that while the insect taxa that respond most directly to the non-native plant diversity are those that rely on those plants as hosts, pollinators, and plant visitors, over time those non-native herbaceous insects support introduced predators and parasites also.
Because of the “invasion debt” associated with that lag, Bonnamour et al. estimate that newly detected insect invasions will increase by 35% worldwide as a result of only recent plant introductions. They differentiate this “invasion debt” from “future invasions”, meaning the actual introduction of additional species resulting from future trade activities.
The model developed by Bonnamour et al. points to the highest numbers of newly introduced insect species occurring in areas with less capacity to deal with bioinvasions. Thus, the Afrotropics are anticipated to receive 869 new insect species, or a 10-fold increase over the number currently known to be established in the region. The Neotropics are projected to be invaded by 809 insect species, also a 10-fold increase. The Indomalayan region will probably detect 776 new insect species, a startling 20-fold increase. In reality, the “invasion debt” might not be quite this severe, since – as Bonnamour et al. note several times – the low numbers of introduced insects currently reported for these tropical regions probably partially reflect limited sampling. They note that already a high proportion of insect species intercepted by biosecurity services on imports arriving from Africa and South America are not yet recorded as established in the exporting regions.
Although both the European Palearctic and Australasia have already received many non-native insect species, their “invasion debt” is relatively high: 417 species for Europe, 317 species for Australasia.
The Neotropics are expected to be the greatest source of insect invasions in the future (904 exported species), followed by the European Palearctic (732 species).
Bonnamour et al. did not include non-native plant species used in agriculture, forestry, or ornamental horticulture. As noted above, these widespread deliberate plantings also affect insect fauna and higher trophic layers.
The greatest number of recorded insect introductions so far are in the Nearctic, Oceania (primarily Hawaii), Europe, and Australasia. While this imbalance is probably caused in part by the significantly limited sampling of non-native insect species in the Asian Palearctic and tropics, it is also true that these regions have received the majority of plant introductions through 1900. This factor has changed in the century since then; many non-native plant species have been recorded in the Afrotropics, Oceania, and Asia.
Eucalyptus plantation in Kwa-Zulu-Natal, South Africa; Kwa-Zulu-Natal Dept. of Transportation
Bonnamour et al. offer several potential explanations for the lag in detecting introduced insects compared to detecting introduced plants. First, it might be necessary for non-native host plants to reach a threshold of abundance before the associated insects are able to establish and spread. Second, reaching that threshold might require repeated introductions of the insect’s host plant species. Third, since only some of the imported plants are transporting insects, repeated imports of host plants might be necessary for the insect to achieve sufficient numbers to establish. Fourth, while their analysis included all non-native insect species, only some insect feeding guilds – herbivores and pollinators – are probably directly facilitated by introduced host plants. Fifth, plant species’ presence tends to be more quickly recorded than insects’ presence. Indeed, MacLaughlin et al. reported a median delay of 80 years between establishment and discovery of plant-feeding Hemiptera. This suggests that the actual time lag between plant and insect establishments might be shorter than the period discussed in Bonnamour et al.
Many insects from the European Palearctic have been introduced to the Nearctic; fewer insects have been introduced in the opposite direction. There is no consensus on the explanation. Thirty years ago Mattson et al. argued that there might be fewer niches for non-native insects in Europe due to the lower host plant diversity in this region caused by the Pleistocene/Holocene glaciations. On the other hand, more plant species from the European Palearctic to the Nearctic than the opposite.
Bonnamour et al. call for further research on:
1) time lags at the scale of individual insect species with their host plants.
2) effects of non-native plants used in agriculture, forestry, or ornamental horticulture.
3) whether time lags between plant and insect invasions vary among taxonomic groups, feeding guilds, or among regions.
4) effect of non-native plant abundance, rather than just species richness, on non-native insect establishment.
Recommendations
Writers about interactions of non-native plant species and insect introductions make a common plea: limit the introduction and spread of non-native plants in order to prevent future invasions of both plants and insects. Bonnamour et al. suggest including the risk of insect introductions in plant invasion risk screening tools. Earlier, the Tallamy and Lalk teams called for ending widespread planting of non-native plants.
USDA Secretary Tom Vilsack
Will policy-makers accept this advice?
I believe that these same interaction of plant host and “pest” introductions presumably applies to pathogens, too. I reiterate my frequent complaint that regulators have not responded to two or more decades of criticism of the failures of the international phytosanitary system re: insect and pathogen introductions via the international nursery trade. Examples include Brasier 2008; Liebhold el. al. 2012; Santini et al. 2013; Roy et al. 2014; Eschen et al. 2015; Jung et al. 2015; Meurisse et al. 2019; O’Hanlon et al. 2021.
As I have said earlier, I appreciate that some scientists are trying to reduce scientific uncertainty about the invasive potential of pathogens native to regions other than North America; I refer here to Jiri Hulcr (see Li et al.), Mech, and Schultz. Many more such studies are needed, addressing potential impacts on a wider variety of North American host trees and shrubs.
The late (& very much lamented!) Gary Lovett of the Cary Institute had advocated halting imports of plants that are congenerics of important North American tree species, in order to minimize the risk that pests that damage those genera will be introduced.
In January I suggested that at the global level we need:
National agricultural agencies, stakeholders, FAO & International Plant Protection Convention (IPPC) should consider amending the IPPC requirement that scientists identify a disease’s causal agents before regulating it. Experience shows that this policy virtually guarantees that pathogens will continue to enter, establish, & damage natural and agricultural environments.
National governments & FAO / IPPC should fund greatly expanded research to identify microbes resident in regions that are important sources of origin for traded plants, vulnerability of hosts in importing countries, and new technologies for detecting pathogens (e.g., molecular tools, volatile organic compounds [VOCs]).
Researchers & agencies should expand international “sentinel plants” networks; incorporate data from forestry plantations, urban plantings, etc. of non-native trees.
NPPOs should adopt regulations that apply the “systems approach” or HACCP programs outlined in ISPM#36. I had discussed these approaches in my Fading Forests III report – link at end of this blog.)
I suggested further that Americans need to
Evaluate the efficacy of current regulations – that is, implementing NAPPRA & Q-37 revision. This evaluation should be based on AQIM data, not port interception data. It should include arthropods, fungal pathogens, oomycetes, bacteria, viruses, nematodes. It should include threats to U.S. tropical islands (Hawai`i, Puerto Rico, Guam, etc.) which are centers of plant endemism.
Apply existing programs (e.g., NAPPRA, Clean Stock Network, post-entry quarantine) to strictly regulate trade in plant taxa most likely to transport pests that threaten our native plants; e.g., plants belonging to genera shared between North American trees & plants on other continents.
Recognize that plant nurseries are incubators for microbial growth, hybridization, and evolution; require nurseries to adopt sanitary operation procedures regardless of whether they sell in inter-state or intra-state commerce
SOURCES
Bonnamour, A., R.E. Blake, A.M. Liebhold, H.F. Nahrung, A. Roques, R.M. Turner, T. Yamanaka, and C. Bertelsmeier. 2023. Historical plant intros predict current insect invasions. PNAS 2023 Vol. 120 No. 24 e2221826120 https://doi.org/10.1073/pnas.2221826120
Burghardt, K. T., D. W. Tallamy, C. Philips, and K. J. Shropshire. 2010. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 1(5):art11. doi:10.1890/ES10-00032.
Lalk, S. J. Hartshorn, and D.R. Coyle. 2021. IAS Woody Plants and Their Effects on Arthropods in the US: Challenges and Opportunities. Annals of the Entomological Society of America, 114(2), 2021, 192–205 doi: 10.1093/aesa/saaa054
Li, Y., C. Bateman, J. Skelton, B. Wang, A. Black, Y-T. Huang, A. Gonzalez, M.A. Jusino, Z.J. Nolen, S. Freeman, Z. Mendel, C-Y. Chen, H-F. Li, M. Kolařík, M. Knížek, J-H. Park, W. Sittichaya, T-H.
Pham, S. Itoo, M. Torii, L. Gao, A.J. Johnson, M. Lur, J. Sun, Z. Zhang, D.C. Adams, J. Hulcr. 2022. Pre-invasion assessment of exotic bark beetle-vectored fungi to detect tree-killing pathogens. https://apsjournals.apsnet.org/doi/full/10.1094/PHYTO-01-21-0041-R
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Liebhold, A.M., T. Yamanaka, A. Roques, S. August, S.L. Chown, E.G. Brockerhoff & P. Pyšek. 2018. Plant diversity drives global patterns of insect invasions. Sci Rep8, 12095 (2018). https://doi.org/10.1038/s41598-018-30605-4
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Mattson, W. J., P. Niemela, I. Millers, and Y. Ingauazo. 1994. Immigrant phytophagous insects on woody plants in the United States and Canada: an annotated list. USDA For. Ser. Gen. Tech. Rep. NC-169, 27 pp.
Mech, A.M., K.A. Thomas, T.D. Marisco, D.A. Herms, C.R. Allen, M.P. Ayres, K.J.K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, and P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol. 2019 Nov; 9(21): 12216-12230.,
Richard, M., D.W. Tallamy and A.B. Mitchell. 2019. Intro plants reduce species interactions. Biol Invasions https://doi.org/10.1007/s10530-018-1876-z
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions.
Tallamy, D.W., D.L. Narango and A.B. Mitchell. 2020. Ecological Entomology (2020), DOI: 10.1111/een.12973 Do NIS plants contribute to insect declines? Conservation Biology DOI: 10.1111/j.1523-1739.2009.01202.x
Uden, D.R, A.M. Mech, N.P. Havill, A.N. Schulz, M.P. Ayres, D.A. Herms, A.M. Hoover, K.J. K. Gandhi, R.A. Hufbauer, A.M. Liebhold, T.D. M., K.F. Raffa, K.A. Thomas, P.C. Tobin, C.R. Allen. 2023. Phylogenetic risk assessment is robust for forecasting the impact of European insects on North American conifers. Ecological Applications. 2023; 33:e2761.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
plane trees in Parco della Montagnola, Bologna, Italy
The CISP blog has had a pause while I travelled in Italy, Slovenia, and Croatia. My purpose was tourism … but I could not forget my interest in invasive species! Most prominent was that global traveller, Ailanthus altissima. I saw lots of the trees in both urban and rural settings. I am unclear about whether some other plant taxa are native v. non-native, e.g., ivy and several species of pines widespread along the Dinaric mountains. Scotch broom Cytisus scoparius was widespread along the Dalmatian coast – but it is native there so not a concern. (Should have been a warning that it would invade other areas with Mediterranean climates!) We were told that the city of Split is even named for the shrub – a shortened/corrupted version of the Greek name for the plant.
Regarding tree-killing pests, I was pleased to see lots of young olive trees along the Dalmatian coast – apparently the Xylella fastidiosa bacterial pathogen that is killing trees in Italy, France, Spain and Portugal is not yet a problem there.
I also saw many of the columnar cedars that I understand are under attack by fungi in the genus Seiridium in Italy. I also saw apparently healthy boxwoods at our hotel near Dubrovnik.
Some of the boxwoods growing in monastery cloisters in Dubrovnik looked as if they have been hit by box tree moth (Cydalima perspectalis) — which is no surprising since the pest was detected in northern Croatia a decade ago.
Dominican monastry in Dubrovnik
Finally, I saw apparently healthy plane trees growing in parks in Bologna; apparently this region has not yet been invaded by the canker stain disease caused by North American fungus Ceratocystis platani. – reported to have caused high levels of mortality in Italy, France, and especially in Greece. (See photo at top of blog) [See Tsopelas, Santini, Wingfield and de Beer. 2017. Canker stain: a Lethal Disease Destroying Iconic Plane Trees. Plant Disease. 2017. 101:645-658. American Phytopathological Society]
I am concerned that the forests at Plitvice National Park are composed of European beech, which is vulnerable to beech leaf disease if it arrives there.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm