The Old Man – a giant ash tree in Wytham Woods; photo from https://theoldmanofwytham.com/2018/11/29/ash-dieback-in-wytham-woods/
I campaign for protecting trees – especially trees growing to their natural capacity in the habitats in which they have evolved. I focus on the threat to these trees from non-native insects and various pathogens (fungi, nematodes …). I have often expressed my distress because others appear to place a low priority on this goal. I have also asked whether protecting trees might be given a higher priority by more decision-makers if they recognize trees’ vitally important role in countering climate change.
For this reason, I have blogged several times about studies examining the role trees play in sequestering carbon — see here & here & here.
A new study demonstrates that protecting large, old trees – almost by definition in their natural environment – is vitally important. Planting new, small, trees is helpful but cannot substitute for the venerable trees.
Calders and colleagues (full citation at the end of the blog; open access!) have used new technology to update assessments of the amount of carbon sequestered in trees. They conducted their study in a temperate hardwood forest – Wytham Woods, a typical broadleaf temperate forest in Oxfordshire, southern Great Britain. [Wytham Woods is also the site of two of the “Inspector Morse” mysteries – “Secret of Bay 5B” and “A Way Through the Woods”.]
They found that these trees sequester 1.77 times more carbon in their above-ground biomass (AGB) than previously believed based on currently-used models.
One consequence of their findings is that countries using the standard assessment method (which was developed by Robert Bunce in 1968) are reporting inaccurate carbon sequestration estimates to the United Nations per the Paris climate accords. (Calders et al. believe that calculations for conifer species are probably more accurate than those for deciduous forests.)
A second consequence is that death of large trees – from whatever cause – will result in greater loss of carbon storage than previously thought.
Old v. New Measurements
The underlying Bunce dataset and algorithm applied in most European biomass estimates were based on a small sample: 200 trees belonging to five taxa growing in one forest area. The models were derived by cutting down trees and weighing them to determine tree biomass. Smaller trees were used because they are easier to process. The scientists then extrapolated the biomass of bigger trees based on the assumption that correlation between tree size and mass is independent of tree size. This assumption has rarely been tested because of the difficulty and expense of carrying out this type of destructive sampling.
The higher estimates of carbon storage in Calders et al. arise in part from the bias towards small trees in calibration of the earlier models. Calders et al. found that trees do not follow a size-invariant scaling relationship, particularly at larger size; it is important to include crown area. Thus, Calders and colleagues calculated a higher sequestration rate for trees in Wytham Woods that fell within the size range used in developing the Bunce allometric model.
In addition, changes in forest management have increased the abundance of larger trees compared to the 1960s when Bunce carried out his study. Indeed, many of the trees in Wytham Woods are nearly twice as large as the trees used in the original calculation of biomass. The median dbh in Bunce (1968) is 8.4 cm; the mean dbh for the TLS dataset (based on a 2015 inventory) is 15.9 cm. The large trees represent a high proportion of the above-ground biomass: 50% of AGB in Wytham Woods was associated with fewer than 7% of the trees (those with dbh greater than 53.1 cm). All these trees were larger than the trees used to calibrate the widely used allometric model.
Calders et al. say that the distribution of tree size (trunk diameter) in Wytham Woods is representative of broadleaved species throughout Great Britain. Basal area had doubled in 40 years from 1974. Thus, the growth trajectory reflected at Wytham Woods – and presumably across Britain – resulted in a net carbon sink of ~1.77tha-1year-1ha in Calder et al’s 3D analysis. This is almost double the ~1tha-1year-1ha derived using the traditional allometric models. .
Methodology
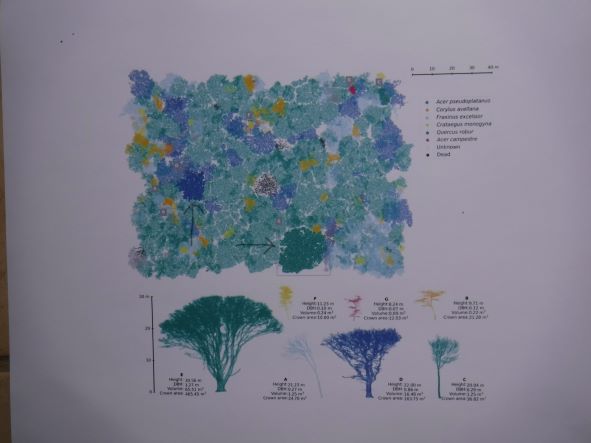
graphic from Calders et al. large maple (green) and oak (blue) trees illustrated by LiDAR images – profiles and location in the forest (indicated by arrows); copyright Ecological Solutions and Evidence
Calder et al. used terrestrial laser scanning (TLS; terrestrial LiDAR) methods & 3-dimensional analysis to derive tree volume and convert this to above-ground biomass (AGB) and carbon sequestration. They scanned 815 live standing trees in Wytham Woods during winter so leaves did not complicate computations. They found:
total volume of these 815 trees was 742.6±3.9m3ha-1.
TLS-derived AGB = 409.9tha-1. This is significantly greater than the 231.9tha-1 resulting from applying the Bunce allometric models.
In sum, 1.77 times more carbon is stored per ha according to this model than carbon values derived through the allometric AGB models developed by Bunce.
Calder et al. describe the threat to European carbon sequestration projections caused by ash dieback. Ash dieback has been spreading across Europe since the 1990s – although the causal agent was not determined until 2006 (Paap et al.). It is killing European ash across the continent. Some of these trees are large – that is, store impressive amounts of carbon. In Wytham Woods specifically, ash dieback threatens some of the largest trees.
Ash dieback disease was first observed in the United Kingdom in 2012; it reached Wytham Woods in 2017. Ash contributed ~13.2% of the biomass carbon sequestration in the study area. However, the species’ presence in all of Wytham Woods might approach ~34%. Ash comprised 75% of seedlings in 2012. Ash is one of three species that contribute >26% of broadleaved tree AGB & carbon for Great Britain as a whole. The British Woodland Trust expects the UK to lose 80% of its ash trees. As a result, Wytham Woods, Britain, and, by extension, a significant amount of European temperate deciduous forests, are likely to become a substantial carbon source in the next decades.
A dead elm tree on Skelston Moor; photo by Walter Baxter; CC BY-SA 2.0
I note that Europe has already lost any sequestration benefits it would have enjoyed from large elm trees due to “Dutch” elm disease. Various Phytophtoras are killing trees in Britain and Ireland.
I recently described threats to plane trees, pines, and other trees across Europe.
I interpret these findings as demonstrating that protecting large trees growing in natural ecosystems is highly important as we try to cope with climate change. This will require determined, sustained, and strategic actions in the face of disturbances predicted to increase as result of changes in climate and the human activities that contribute to climate change – e.g., overexploitation of natural resources, conversion of natural systems to human use, shipping goods around the globe, …
Calders and colleagues say we cannot afford to lose substantial reservoirs of carbon currently sequestered in temperate forests. Such forests currently account for ~14% of global forest carbon stocks in their biomass and soil. Their importance is growing because of widespread deforestation in the tropics.
What is To Be Done? (to cite Lenin)
Calders and colleagues call for several actions to address potential biases in biomass carbon estimates and drastically improve estimates of forest biomass:
(i) Research to improve knowledge about carbon sequestration levels in trees. This will require
a) greater sampling using such nondestructive methods as TLS to estimate AGB of a wider variety of forest types,
b) improved understanding of wood density, and
c) properly testing the fundamental assumption of size dependency in allometric models.
(ii) Develop empirical models of AGB that do not assume size invariance. This might require. This implies more destructive harvesting to obtain data from a variety of forest compositions, locations, etc,
(iii) Establish a biomass reference network of permanent sample plots specifically designed for estimating AGB. The improved data can then be fed into satellite-derived biomass estimates, which are likely to become the de facto standard for assessing the state and change of forest AGB at large scales. The GEO-TREES database can help. It aims to build on existing long-term ecological plot networks, by including TLS, airborne laser scanning & other ancillary data (including harvest measurements) to specifically allow for upscaling of AGB & development of new empirical models.
(iv) Ensure much better traceability in the use of allometric models. If applying a model to a site at several removes from the original data, e.g., published allometric models, clearly identify where and when the underpinning data were collected, the number and size range of trees from which models were derived, and clarify any assumptions regarding environmental conditions, wood density etc. Database initiatives such as GlobAllomeTree can help.
North American Situation
remains of Michigan’s champion green ash
A study in 2019 (Fei et al. 2019; full citation at end of the blog) has already estimated that 41% of total live (woody) biomass in forests of the “lower 48” states is at risk from the most damaging of introduced pests. The greatest biomass loss was caused by emerald ash borer, Dutch elm disease, beech bark disease, and hemlock woolly adelgid. Before arrival of these non-native pests, mature ash, elms, beech and hemlock were large – providing significant storage of carbon (and other ecosystem services). A complication is that elms and beech, at least, began dying decades before the underlying (Forest Inventory and Analysis; FIA) data began to be collected. Consequently, the reported mortality rates underestimate the actual loss in biomass associated with these pests.
Did Fei et al. rely on biomass estimates based on measurements and algorithms now questioned by Calders et al.? One of the co-authors, Dr. Randall Morin, has told me that USFS scientists are shifting to new models that will result in a slight bump in overall biomass for the U.S. largely because of increased recognition of the biomass in crowns and limbs. However, the new models are based partly on a felled-tree study, so I wonder if they will have similar issues.
Certainly in some situations that threat posed by non-native pests is not yet being adequately incorporated. Badgley et al. (2022) analyzed the California cap-and-trade program to determine whether forest projects enrolled under its provisions can provide sufficiently permanent carbon sequestration. They determined that sequestration losses tied to mortality of one tree species (tanoak; Notholithocarpus densiflorus) due to one disease – sudden oak death – would fully deplete the “buffer pool” set aside to compensate for losses due to disease and insect infestations. This leaves the program unable to provide the promised benefits in carbon sequestration. SOD continues to spread and tanoaks (and other tree species) to die. California along is home to other tree-killing pathogens and insects, e.g., white pine blister rust, Port-Orford cedar root disease, Fusarium dieback, goldspotted oak borer …
California live oak killed by GSOB; photo by F.T. Campbell
Furthermore, the program allows enrollment of forests across the United States, so the multiple pests threatening ash, hemlocks, oaks, and other tree taxa across North America must also be accommodated. I have not even mentioned the likelihood that additional tree-killing pests will be introduced in the future.
How can scientists enhance the credibility of well-intentioned efforts to incorporate forest conservation into strategies aimed at mitigating climate change?
[A separate study by Oxford University has estimated that 2 billion tonnes of CO2 are removed from the atmosphere every year – 99% of it by trees. They point out that this is not sufficient to help Earth avoid temperatures rising above Paris-set levels. See an article by Lottie Limb, Reuters, published 19 January 2023 (sorry – I don’t have a direct link).]
SOURCES
Badgley, G., Chay, F., Chegwidden, O.S., Hamman, J.J., Freeman J. and Cullenward, D. 2022. Calif’s forest carbon offsets buffer pool is severely undercapitalized. Front. For. Glob. Change 5:930426. doi: 10.3389/ffgc.2022.930426
Bunce, R. G. H. (1968). Biomass and production of trees in a mixed deciduous woodland: I. Girth and height as parameters for the estimation of tree dry weight. Journal of Ecology, 56, 759–775.
Calders, K., H. Verbeeck, A. Burt, N. Origo, J. Nightingale, Y. Malhi, P. Wilkes, P. Raumonen, R.G.H. Bunce, M. Disney. Laser scanning reveals potential underestimation of biomass carbon in temperate forest. Ecol Solut Evid. 2022;3:e12197. wileyonlinelibrary.com/journal/eso3 open access!
Paap, T., M.J. Wingfield, T.I. Burgess, J.R.U. Wilson, D.M. Richardson, A. Santini. 2022. Invasion Frameworks: a Forest Pathogen Perspective. FOREST PATHOLOGY Current Forestry Reports https://doi.org/10.1007/s40725-021-00157-4
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Spruce budworm (Choristoneura fumiferana); photo by Jerald E. Dewey, USFS; via Bugwood; populations of several forest birds, including Cape May, Tennessee and Bay-Breasted warblers, become more numerous during budworm outbreaks
Now that Drs. Ziska and Aucott have educated us about the strong impact atmospheric CO2 can have on both plants and phytopagous insects, I have asked the experts whether these interactions have been incorporated in the models scientists are using to forecast pest activity in American forests as the climate changes.
The answer is no.
bay-breasted warbler; photograph by Dave Inman at Presque Isle State Park, PA; via Flickr
Dr. Bethany A. Bradley, Co-Director, Northeast Climate Adaptation Science Center at the University of Massachusetts, says empirical models of species range shifts typically only use climate and sometimes other environmental factors (like soils or topography) as predictors of potential geography. Inclusion of demographic processes like how plant growth is affected by more or less water, CO2, competition with other plants etc. would require a lot of data. It is currently impossible since there are tens of thousands of plant species interacting in the forests of eastern North America – and perhaps these factors have been analysed for only a hundred of them.
Mike Aucott points to the same difficulty: inclusion of CO2 in models of the future populations of specific plants would be difficult since the impacts vary from species to species and are compounded by other factors such as soil nitrogen levels, moisture levels, temperature, presence of competing plants, etc.
Regarding insects, Dr. Aucott thinks it is clear that some orders, such as Lepidoptera, don’t fare as well when feeding on plants grown under elevated CO2. He is not aware of efforts to model impacts of high CO2 on specific insects or even orders or feeding guilds.
juniper geometer (inchworm); Dr. Tallamy says inchworms are hairless & good tasting – so sought by birds
Dr. Ziska concurs about the difficulties. Dr. Ziska asks why there is so little funding to study these issues, especially given their probable impact on human food supplies and health – as described in his blog and an opinion piece published in Scientific American two years ago.
I hope that scientists, decision-makers, readers of this blog … maybe even the media! – take into consideration these complexities, even if they cannot be defined.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – [but do not address climate or CO2 aspects] review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Wildfire: one of the widely recognized results of climate change (The Pioneer Fire located in the Boise National Forest near Idaho City, ID began on Jul. 18, 2016 and the cause is under investigation. The Pioneer Fire has consumed 96,469 acres. U.S. Forest Service photo. Original public domain image from Flickr)
A guest blog by Michael Aucott. Mike is a retired research scientist of the NJ Department of Environmental Protection. He has also taught chemistry at the College of New Jersey. He is currently a member of the NJDEP Science Advisory Board Standing Committee on Climate and Atmospheric Sciences, and on the board of directors of the PA/NJ Chapter of the American Chestnut Foundation. If you wish to contact Mike, use the contact button on this website. You MUST include your email address; it is not recorded automatically.
Two major perturbations affect Earth and its living systems, climate change and habitat disruptions. Emerging data show that these are more closely related than previously realized; they are connected by carbon dioxide, CO2.
Climate change basics: the physics
Climate change concerns have focused on the alteration of weather and climate due to the increase in atmospheric concentrations of greenhouse gases, primarily carbon dioxide, CO2. The impact of CO2 on climate has been understood for at least 120 years. In 1896 the Nobel-Prize-winning Swedish chemist Svante Arhennius published calculations demonstrating that human emission of CO2, when combined with the positive feedback effects of water vapor, would warm the Earth (Arhennius, 1896). His equation, ΔF = α ln(C/C0), relates the change in climate “forcing” (the degree to which temperature change is forced) to the ratio of the concentration of CO2 currently in the atmosphere (C) to a previous concentration (C0). This equation is still in use today. Arhennius estimated that a doubling of CO2 would warm the Earth by about 4 degrees C. This estimate is not far off from current estimates based on much more elaborate calculations.
This warming impact is caused by the physics of CO2, water vapor, and other “greenhouse” gases. Infrared radiation causes the CO2, water, and other greenhouse gas molecules to vibrate, leading to the absorption of the energy carried by that radiation. Much of the solar energy coming from the sun is not in the infrared frequency range, so it passes through the atmosphere without being absorbed. However, when this energy is absorbed by the surfaces of the Earth and its biota, and is re-radiated as infrared radiation, it is then absorbed by greenhouse gases, warming the planet.
The amount of water vapor in the atmosphere is directly related to the atmosphere’s temperature: warm air holds more water vapor. Human activity hasn’t directly changed the concentration of water vapor in the atmosphere significantly. But by burning fossil fuels, humans have dramatically increased the atmospheric concentration of CO2 and in so doing, also indirectly increased the concentration of water vapor. Just as Arhennius predicted over 120 years ago, this increase in CO2 is warming the Earth.
Ramifications of this warming include increased heat episodes, the intensification of the hydrological cycle (greater frequency of both heavy precipitation events and of droughts), sea level rise due to the melting of land-based glaciers and the thermal expansion of ocean water, and, almost certainly, intensification of storms and an increase in extreme weather. These climate-warming- based perturbations have the potential to influence the functioning of Earth’s biota in many deleterious ways, and clearly can be associated with the many facets of habitat disruption.
Climate change amplifications: the chemistry
But there’s another aspect of CO2 that may be more important insofar as habitat disruption is concerned and that has been largely ignored: chemistry. CO2 is a trace gas as far as we humans and other animals are concerned, unnoticed by our bodies in normal life. But to plants it is a vital food. It is taken up by plants as an essential input to photosynthesis. In this chemical reaction, using the energy of sunlight, plants combine CO2 and water vapor to make oxygen and carbohydrates, represented with a generic formula of CH2O, according to the equation CO2 + H2O → CH2O + O2. Without this reaction, life as we know it would not exist.
The atmospheric concentration of CO2 has varied over time; some 50 million years ago it was considerably higher than today. However, for at least the last three million years, the concentration of CO2 has been in the range of 280 ppm. Over these millions of years biota have adapted to this concentration. But within the last 300 years, one ten thousandth of this period – a blink of an eye in the geological or evolutionary time scale – the concentration of CO2 has shot up to 420 ppm, a 50% increase, due to humanity’s burning of fossil fuels and forests.
Imagine what might happen to a person who had been on a tight dietary budget for most of his or her life but suddenly got access to 50% more carbohydrates, but no more protein or minerals? We could expect major changes in the metabolism of that person. This dramatic change is what has, in effect, happened to the whole of life on Earth. Our planet’s primary biota, plants, now suddenly have the opportunity to gorge on CO2. But their access to other important substances such as nitrogen has not changed. Evidence is accumulating that CO2 at its elevated level of 420 ppm is not, as has been proclaimed by some, a healthy influence but is instead throwing Earth’s ecosystem into disarray.
Much of the recent experimental evidence on the impacts of enriched atmospheric CO2 has been assembled by Lewis Ziska and presented in his new book, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It (Ziska, 2022; see full citation at the end of the blog). The findings documented in this book reveal a variety of impacts of elevated CO2. These impacts include stimulation of growth of invasive plants, decreases in the nutrient content of major crops, and changes in plants’ production of insecticidal, allergenic, and other compounds. The changing chemistry of plants may be contributing to a major die-off of insects and insect-eating animals including birds. Below are some details.
Habitat Disruptions: Stimulation of Invasive Plants
The generally accepted explanation for why some plants are invasive is that they have been introduced to new regions where their historic predators and parasites aren’t present. Without these drags on their growth, they have flourished. That some alien plants are not browsed by white-tailed deer contributes significantly to their invasiveness in Eastern North America. Other factors are clearly involved as well, including changes in the temperature regime and the availability of water and other resources such as nitrogen.
But elevated CO2 is also a factor. In recent years, techniques for measuring responses of organisms in situ under elevated CO2 conditions have been developed, making possible investigations of the impacts of CO2 concentrations that could exist in the future under otherwise relatively realistic conditions. What the actual atmospheric CO2 concentration will be in 2050 or 2100 is difficult to predict; it depends on what humanity does to control emissions. Various scenarios suggest that levels could exceed 500 ppm by 2050 and might exceed 1000 ppm by 2100 (Tollefson, 2020).
cheatgrass; photo by Jaepil Cho
One study found that the invasive weed Canada thistle, Cirsium arvense, responds more strongly to elevated CO2 than soybean, a crop that it often plagues. In a high CO2 environment, this weed’s root system grows strongly enough to make it significantly more resistant to herbicides (Ziska, et al., 2004) (Ziska, 2010). The highly invasive and dangerously flammable cheatgrass (Bromus tectorum), nicknamed “grassoline” by the U.S. Forest Service because of its propensity to intensify wildfires, also responds strongly to elevated CO2 (Ziska, et al., 2005). Also found to be boosted by enriched CO2 is yellow star-thistle (Centaurea solstitialis), considered one of California’s worst weeds. In one study (Dukes, et al., 2011) this plant grew 600% larger in elevated CO2 relative to ambient, while native plants responded much less strongly or not at all. Japanese honeysuckle, Lonicera japonica, which plagues many areas in the U.S., was found to increase in biomass by 135% at a CO2 concentration of 675 ppm while a similar native plant, coral honeysuckle (Lonicera sempervirens) increased by only 40% (Sasek & Strain, 1991). In a field study also involving Japanese honeysuckle (Belote, et al., 2004), researchers found that its above ground net production (ANPP) approximately tripled under enriched CO2 while other plants in the trial showed showed lesser increases or actual decreases.
Other plants have been found to be selectively encouraged by enriched CO2 including cherry laurel (Prunus laurocerasus), invasive in the Pacific Northwest U.S. and the U.K., (Hattenschwiler & Korner, 2002); dalmation toadflax (Linaria dalmatica), invasive in much of North America (Blumenthal, et al., 2013); honey mesquite (Prosopis glandulosa) , invasive in Australia and parts of Africa (Polley, et al., 1996); and kudzu (Pueraria lobata), which afflicts the Southeast U.S. (Sasek & Strain, 1988). Three plants invasive in China or Southeast Asia, American rope (Mikania micrantha), Creeping oxeye (Wedelia trilobata), and a morning glory species (Ipomoea cairica), were found to produce 70.3% greater biomass when grown at a CO2 concentration of 700 ppm while three corresponding indigenous plants Paederia scandens, Wedelia chinensis and Ipomoea pescaprae, produced only 30.5% more biomass (Song, et al., 2009).
yellow star thistle; photo by Eugene Zelenko
The list goes on of studies showing increased growth of some plants under enriched CO2 conditions. As more in situ investigations are undertaken, it seems likely it will become clearer that today’s enriched level of CO2 is helping some plants to become invasive.
Complexities and contradictory findings exist however. Not all plants are stimulated by enriched CO2. An important difference in the response to higher levels of CO2 is whether a plant has a C3 or a C4 photosynthetic mechanism. C4 plants contain a biochemical pump that concentrates CO2, making them more adapted to low CO2 conditions (Hager, et al., 2016). At current levels of CO2, such plants’ need for CO2 is easily met. C3 plants do not have this CO2 concentrating ability, and so higher levels boost their growth. In a broad meta-analysis of literature, the average response to elevated CO2 of 365 C3 plant species and 37 C4 plant species was noted; the response was significantly increased in C3 species but was unchanged in C4 species (Robinson, et al. 2012). One striking example of such a difference was observed in the field study noted above (Belote, et al., 2004), wherein researchers found that Japanese honeysuckle (a C3 plant) was significantly encouraged by elevated CO2 relative to other plants at the same locale. The same study found that another aggressive invader, Japanese stiltgrass (Microstegium vimineum), a C4 plant, was unaffected or even slightly inhibited relative to competing plants’ growth by elevated CO2.
One finding is widespread; most plants studied under enriched CO2 regimes show an increase in biomass and evince a higher ratio of carbon to nitrogen (C/N ratio) in their tissues and an overall decline in nitrogen concentrations than when grown under ambient conditions. Since nitrogen is a key component of protein, this change can be expected to lead to lowered protein content of critical food crops. Some impacts of this change are already well underway because of today’s elevated CO2 concentration. Changes since 1850 in the C/N ratio and in the estimated protein content of an important plant product, pollen, were discovered in a striking study by Lewis Ziska and colleagues (Ziska, et al., 2016). Using archived museum samples, these researchers determined the nitrogen content of pollen of Solidago canadensis (Canada goldenrod) going back to the 1850s. They estimated that the protein content of goldenrod pollen, a vital nutrient for North American bees, has declined in inverse proportion to the rise in atmospheric CO2, dropping from a concentration of approximately 18% in the mid-1800s to approximately 12% today. They pointed out that it is possible that bees are now unable to provide sufficient protein and other nutrients to larvae, and that one of the main reasons for bee declines is malnutrition caused by enriched atmospheric CO2. Other studies also indicate that elevated CO2 could cause lower nitrogen concentrations in plants and lead to less proteinaceous plant parts, including pollen, being available to plant-feeding insects (Hall, et al., 2005; Knepp, et al., 2007).
bumblebee on goldenrod; photo by Keila
The changing C/N ratio is almost certainly already affecting the human food supply. As documented in an extensive review of published findings (Soares, et al., 2019), elevated CO2 has a considerable impact on the accumulation of minerals and protein in plants, with many plant species showing declines in both quality and quantity of key nutrients. These changes have worrisome implications for human nutrition and may already be responsible for increasing incidences of dietary deficiency in some areas. Lewis Ziska discusses the likely impact of rising CO2 on the future human food supply in his recent post. A number of studies showing declines in protein and also other nutrients such as zinc in food crops important to humanity are also described in Ziska’s new book, Greenhouse Planet, noted above.
Habitat Disruptions: Other Changes in Plant Chemistry
Other changes in plants besides nutritional content may be driven by enriched CO2. Plants produce a variety of secondary metabolites. Most plants use the C3 mechanism; with 50% more available of a key input, some changes in these plants’ production of such chemicals can be expected. Some changes have been observed. Mohan et al. (2006) report that enriched CO2 in an intact forest system increased water use efficiency, growth, and population biomass of poison ivy (Toxicodendron radicans) and that high-CO2 plants also produced a more toxic form of the allergenic compound urushiol.
Quercus chapmanii; photo by Mary Keim at Seminole State Forest, Florida
Many of the phytochemicals plants produce function as defenses against insect predation, and changes in such production have been found to impact herbivore feeding. For example, Landosky and Karowe (2014) suggest that specialist herbivores may have to contend with more effective chemical defenses by plants under elevated CO2. Hall, et al. (2005), in a study involving several oak and one legume species in a scrub oak ecosystem in Florida (see photo above), found that 700 ppm CO2 levels led to decreased damage to plants by four of six insect groups investigated. They did not see increases in plants’ production of carbon-based secondary metabolites, including total phenolic compounds, condensed tannins, hydrolyzable tannins, cellulose, hemicellulose, and lignin however. They concluded that the primary driver of decreased insect predation under elevated CO2 was lower overall plant nitrogen levels. They stated that the decline of nitrogen levels in foliage under elevated CO2 indicated lower foliar quality and hypothesized that the reductions in insect feeding stemmed from the combined effects of nutrient limitation and increases in parasitism and predation on the nutrient-constrained insects. They further stated that although insects try to compensate for lower nutrient content of leaves by eating more, they did not see an increased portion of damaged leaves in their study. These researchers did not report measurements of terpenoid compounds however, which are reported to represent the largest class of secondary metabolites (Wikipedia, 2022). In another study (Hall, et al., 2005a) found that concentrations of condensed tannins were higher in oak leaf litter under elevated CO2, which suggests that enhanced production of insecticidal compounds or other changes to plant tissues could affect not only insects that consume living plant tissue, but also detritivores.
Robinson et al. (2012) also investigated plants’ production of secondary metabolites in their literature review. Looking at all plant groups, they found that under elevated CO2 the production of nitrogen-based secondary metabolites (e.g., alkaloids, cyanogenic glycosides, and glucosinolates) decreased by 16% while the carbon-based secondary metabolites total phenolics, condensed tannins, and flavonoids increased by 19%, 22%, and 27% respectively. Another carbon-based metabolite, terpenoids, declined by 13%. They further divided plants into grasses, shrubs, herbs/forbs, and trees and found differing responses to elevated CO2. Trees, for example, showed increased production of total glycosides and total phenolics, little change in production of total flavonoids, and a decline in the production of total terpenes. Like Hall et al., (2005), Robinson et al. found a strong and significant decrease in nitrogen concentrations under elevated CO2 for C3 plants. A decrease did not show up for C4 plants.
In addition to chemical defenses, plants have physical characteristics such as surface waxes, trichomes, secretory canals, and tissue toughness-enhancing substances such as lignin. All of these features can reduce the edibility of plants for arthropod herbivores. Robinson et al. (2012) found consistent responses to these characteristics under elevated CO2; leaf toughness and specific leaf weight increased by 11% and 18%, respectively, while specific leaf area did not show a significant change. They concluded that there is an increase in general “toughness” of leaves under elevated CO2. As did Hall et al., (2005), Robinson et al. concluded that elevated CO2 will induce changes in plant chemistry, physiology, and morphology that are likely to impact the nutritional quality of host plants for insect herbivores.
Habitat Disruptions: Changes in Plant Chemistry and Insect Decline
Numerous studies have documented a recent and dramatic decline in insect populations and discussed the probable cascading impacts of such declines through the food chain, affecting spiders, lizards, birds, and other organisms (Samways, et al., 2020; Cardoso, et al., 2020; Sánchez-Bayoa & Wyckhuys, 2019; Tallamy & Shriver, 2021). It has been argued that the main drivers of insect species declines are habitat loss and conversion to intensive agriculture and urbanization; pollution, mainly by synthetic pesticides and fertilizers; biological factors, including pathogens and introduced species; and climate change.
But a puzzling aspect is that some insect declines have been observed in nature preserves (Vogel, 2017) that presumably are not greatly affected by most of the above drivers. One example is a study spanning 27 years that found a 76% decline in flying insect biomass at several of Germany’s protected areas subject to rather low levels of human disturbance (Hallmann et al., 2017). Another study in rainforests of Puerto Rico, apparently not subject to influences such as light pollution, habitat loss, pesticides, or agriculture, reported biomass losses between 98% and 78% for ground-foraging and canopy-dwelling arthropods over a 36-year period, (Lister and Garcia, 2018). This leaves climate change as the likely culprit. But although the varied impacts of climate change, including heat episodes, drought, and other episodes of extreme weather could impact insect populations in remote as well as populated areas, the trends observed appear to far exceed the magnitude of such climate-related disturbances over the last several decades.
tent caterpillars; Shiela Brown, Public Domain Pics
Another puzzling aspect is that not all insect orders or feeding guilds seem to be equally affected. Sanchez-Bayoa & Wychuys (2019) whose article reports on a review of 73 historical reports, state that Lepidoptera, Hymenoptera and dung beetles (Coleoptera) appear to be the taxa most affected in terrestrial environments. Robinson et al. (2012) found that phloem feeders such as Homoptera respond positively to elevated CO2 while foliage feeders/Lepidoptera respond negatively. Lepidoptera were among the most impacted; relative growth rate, fecundity, and abundance all declined under high CO2 conditions, while relative consumption rate, total consumption, and development time all increased.
Most Lepidoptera are herbivorous, feeding in their larval stage, caterpillars, on plants. Caterpillars are key components of the terrestrial ecology; in most forests of the world, caterpillars consume more living leaves than all other animals combined (Janzen, 1988). Insect herbivores such as caterpillars are near the hub of most terrestrial food webs, comprising essential food for insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears (Burghardt et al., 2010). At least 310 North American bird species are known to feed extensively on caterpillars, and the majority of terrestrial birds rely on insects during reproduction and other nutrient-limited periods in the annual cycle (Narango, Tallamy & Marra, 2018). Caterpillars and moths are the largest component of nestling diets in hundreds of species of migrant bird species (Tallamy & Shriver, 2021); they are among the “little things that run the world” (Wilson, 1987).
Carolina chickadee; one of the birds Dr. Tallamy focuses on because it feeds its young on caterpillars; photo by Dan Pancamo; through Wikimedia
Habitat Disruptions and Climate Change: Connected by CO2
The apparently heightened rate of decline of insect herbivores such as caterpillars compared to some other insects, and the findings that many declines have been observed in areas relatively unimpacted by direct human influences such as light pollution, pesticides, and land-use change, point to the likelihood of a broad, perhaps ubiquitous, cause. Climate change is such a broad cause. Even broader and more ubiquitous is the main driver of climate change, CO2. Every plant in the world is constantly bathed in an enriched concentration of this gas. A conclusion seems likely: CO2 is not only causing global warming and climate change but is also affecting life on this planet in ways that, while still poorly understood, are already reducing the nutritive value of food crops, may be a significant cause of the spread of invasive plants, and may be the main driver of insect declines and the cascading impacts of such declines on insect-eating animals such as birds.
What to do?
To mitigate climate change and, as argued here, to mitigate habitat disruption, the steady rise in the atmosphere’s burden of CO2 must be halted, and then steps must be taken to lower the current concentration to a healthier level. These are not hopeless tasks. Although what has been a relentless rise in CO2 emissions at the global level continues, increases have slowed and even stopped in some parts of the world. Accelerating the development of low- and zero-carbon energy sources and encouraging energy conservation, as will be done through the U.S.’s Inflation Reduction Act, will further this progress.
More will be needed. Putting a significant and steadily increasing price on the carbon in fossil fuels is arguably the most important next step. Fossil fuels enjoy a free ride. The byproduct of their combustion, CO2, is dumped with little or no restrictions into the world’s atmosphere. A price on carbon would end this inequity. There are ways this could be done in a revenue-neutral (“fee and rebate”) manner that would avoid harm to economies and those with low- and moderate-incomes. A major step forward in pricing carbon by the European Union, a carbon border adjustment mechanism, is close to implementation. For more on this and other developments in cutting CO2 emissions, see the analyses and insights of the Carbon Tax Center and learn more about actions you can take to influence governments with Citizens’ Climate Lobby.
Not discussed here, but another stark example of habitat disruption is the increasing acidification of the world’s oceans caused by the dissolution of atmospheric CO2 in the waters. The ocean’s average pH has dropped from 8.2 to 8.1 within the last 200 years. Because pH is a logarithmic scale, this represents an increase in hydrogen ion concentration of over 25%, a change that is already threatening some marine creatures. More on this is available from many sources; e.g., Kolbert (2014).
References
Arhennius, Svante, 1896, On the Influence of Carbonic Acid in the Air upon the Temperature on the Ground, Philosophical Magazine and Journal of Science, 41, 237-276.
Belote, R., J. Weltzin, and R. Norby, 2004, Response of an Understory Plant Community to Elevated [CO2] Depends on Differential Responses of Dominant Invasive Species and Is Mediated by Soil Water Availability, New Phytologist 161, 827-835.
Blumenthal, D., V. Resco, J. Morgan, D. Williams, D. LeCain, E. Hardy, E. Pendall, and E. Bladyka, 2013, Invasive Forb Benefits from Water Savings by Native Plants and Carbon Fertilization Under Elevated CO2 and Warming, New Phytologist 200, 1156-1165.
Burghardt, Karin T., D. W. Tallamy, C. Philips, and K. Shropshire, 2010, Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities, Ecosphere 1: 1-22.
Dukes, J., N. Chiariello, S. Loarie, and C. Field, 2011, Strong Response of an Invasive Plant Species (Centaurea solstitialis L.) to Global Environmental Changes, Ecological Applications 21, 1887-1894.
Hall, M., P. Stiling, B. Hungate, B. Drake, and M. Hunter, 2005a, Effects of elevated CO2 and herbivore damage on litter quality in a scrub oak ecosystem, Journal of Chemical Ecology, 31, 2343-2356.
Hall, M., P. Stiling, D. Moon, B. Drake, and M. Hunter, 2005, Effects of elevated CO2 of foliar quality and herbivore damage in a scrub oak ecosystem. Journal of Chemical Ecology 31, 267-286.
Hallmann, C.A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., Stenmans, W., Müller, A., Sumser, H., Hörren, T., Goulson, D., de Kroon, H., 2017, More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12, e0185809.
Hattenschwiler, S. and C. Korner, 2003, Does Elevated CO2 Facilitate Naturalization of the Non-indegenous Prunus laurocerasus in Swiss Temperate Forests?, Functional Ecology 17, 778-785.
Janzen, Daniel H., 1988, Ecological Characterization of a Costa Rican Dry Forest Caterpillar Fauna, Biotropica, 20, 120-135.
Knepp, R., J. Hamilton, A. Zangeri, M. Berenbaum, and E. Delucia, 2007, Foliage of oaks grown under elevated CO2 reduces performance of Antherae Polyphemus (Lepidoptera: Saturnidae), Environmental Entomology 36, 609-617.
Kolbert, E., 2014, The Sixth Extinction, Henry Holt & Co., NY
Landosky, J., and D. Karowe, 2014, Will chemical defenses become more effective against specialist herbivores under elevated CO2? Global Change Biology, 20, 3159–3176.
Lister, B., and A. Garcia, 2018, Climate-driven declines in arthropod abundance restructure a rainforest food web, PNAS 115, E10397–E10406
Mohan, J., L. Ziska, W. Schlesinger, R. Thomas, R. Sicher, K. George, and J. Clark, 2006, Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2. PNAS 103, 9086-9089.
Narango, D., D. Tallamy, and P. Marra, 2018, Nonnative plants reduce population growth of an insectivorous bird, PNAS 115: 11549–11554.
Polley, H., H. Johnson, H. Mayeux, C. Tischler, and D. Brown, 1996, Carbon Dioxide Enrichment Improves Growth, Water Relations, and Survival of Droughted Honey Mesquite (Prosopis glandulosa) Seedlings, Tree Physiology, 16, 817-823.
Robinson, E., G. Ryan, and J. Newman, 2012, A meta-analytical review of the effects of elevated CO2 on plant-arthropod interactions highlights the importance of interacting environmental and biological variables, New Phytologist 194, 321-336.
Sánchez-Bayoa, F. and K. Wyckhuys, 2019, Worldwide decline of the entomofauna: A review of its drivers, Biological Conservation 232, 8-27.
Sasek, T. and B. Strain, 1988, Effects of Carbon Dioxide Enrichment on the Growth and Morphology of Kudzu (Puerarialobata), Weed Science 36, 28 – 36, DOI: https://doi.org/10.1017/S0043174500074415
Sasek, T. and B. Strain, 1991, Effects of CO2 Enrichment on the Growth and Morphology of a Native and Introduced Honeysuckle Vine, American Journal of Botany 78, 69-75.
Soares, J., C. Santos, S Carvalho, M Pintado, and M. Vasconceios, 2019, Preserving the nutritional quality of crop plants under a changing climate: importance and strategies. Plant and Soil 443, 1-26. https://doi.org/10.1007/s11104-019-04229-0
Song, L., J. Wu, C. Li, F. Li, S. Peng, and B. Chen, 2009, Different responses of invasive and native species to elevated CO2 concentration, Acta Oecologica 35, 128-135.
Tallamy, D. and W.G. Shriver, 2021, Are declines in insects and insectivorous birds related? Ornithological Applications 123: 1–8, DOI: 10.1093/ornithapp/duaa059
Vogel, G., 2017, Where Have All the Insects Gone? Surveys in German nature preserves point to a dramatic decline in insect biomass. Key members of ecosystems may be slipping away, Science 356, 376-379.
Wilson, Edward O., 1987, The Little Things That Run the World (The Importance and Conservation of Invertebrates), Conservation Biology, 1, 344-346.
Ziska, L., 2010, Elevated Carbon Dioxide Alters Chemical Management of Canada Thistle in No-till Soybean, Field Crops Research 119, 299-303.
Ziska, L., Shaun Falukner, and John Lydon, 2004, Changes in biomass and root: Shoot Ratio of Field-grown Canada Thistle (Cirsium arvense), a Noxious, Invasive Wed, with Elevated CO2: Implications for Control with Glyphosate, Weed Science 52, 584-588.
Ziska, L.H., J.S. Pettis, J. Edwards, J.E. Hancock, M.B. Tomecek, A. Clark, J.S. Dukes, I. Loladze, and H.W. Polley, 2016, Rising atmospheric CO2 is reducing the protein concentration of a floral pollen source essential for North American bees, Proc. R. Soc. B, 283, 20160414, http://dx.doi.org/10.1098/rspb.2016.0414
Ziska, Lewis, 2022, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It, Columbia University Press, NY.
Guest blog by Lewis Ziska, Associate Professor, Environmental Health Sciences at the Columbia University
[Dr. Ziska has spent his career analyzing the impacts of CO2 and climate change on plants – and therefore on people. He served as Project Leader for global climate change at the International Rice Research Institute; then spent 24 years at the USDA’s Agricultural Research Service, where he worked primarily on documenting the impact of climate change and rising carbon dioxide levels on: Crop selection improves production; Climate and agronomic pests, including chemical management; Climate, plant biology and public health impacts on food security with a focus on nutrition and pesticide use.]
No question you’ve heard the term, “Climate Change” or “Global Warming”, or my personal favorite, “Global Weirding”. The consequences are talked and discussed in the media—as they should be—but often the media, like many Americans, is focus challenged. Or in more polite terms, they have the attention span of a hummingbird on crack. Which is to say, that simple physical consequences, like sea level rise (heat melts ice!), and stranded animals on ice (Poor polar bear!), or intense storms (newscaster whipped about in the rain, yelling to be understood) are repeated, over and over again. Understandable, makes for good TV.
But it also makes you feel separate from what is happening, these consequences of climate change are to the “other”. I don’t live near the ocean, I don’t interact with polar bears; sure we have storms, but I live in the Midwest, in one of those states that begins with a vowel. Shoot, I commute to work, try and make ends meet, I’m not some damn tree hugger. Why should I care?
To understand why, you need a bit more background, some science that isn’t always available on TV or social media when it comes to global weirding.
First, while you may not be a tree hugger, you do, in fact, interact with nature. Several times. Every day. We call those times, “breakfast”, “lunch” and “dinner”.
dinner; photo by davylin via Flickr
You depend on nature for food. And clothing. And paper. And medicine. And oxygen. And construction materials (wood), and many, many other things. So, if nature gets hinky, and the climate becomes uncertain, it might be worth your while to think about climate change, or global weirding, in a different light. What I want to do here then, is to illuminate two examples that I hope will help you see why climate could affect you, directly and significantly.
Let’s begin with plants. Those green living things that comprise the bulk of the natural world (literally, if you were to weigh the natural world, 97% would be plants, 3% animals). Then let’s look at them through two different lenses—how will climate weirding alter your food; shoot, how will it alter the air that I breathe?
Let’s start with a basic food, rice. Obviously you don’t want to mess with its production, or its nutritional quality. But that is exactly what global weirding is doing.
Rice has flowers. Not big showy ones, but flowers none the less—ones that get fertilized with pollen, and seed is produced. The seed that feeds some two billion people– or about a quarter of the earth’s population.
Like all living things, plants are heat sensitive, and for rice, and many crop plants, the degree of sensitivity varies, depending on the part of the plant in question. Take a look at the table. The crops that are listed, including rice, are the core of what the world eats. Now notice the difference in temperature sensitivity. Vegetative parts of the plant, leaves and stems, are reasonably tolerant of higher temperatures, but flowers are not. Pollen, the plant equivalent of animal sperm, is highly temperature sensitive, and if the temperatures get into the high 90s (37-38oC), they become deformed, and the rice plant doesn’t produce seeds. Same for a number of plants, ones necessary to feed 8 billion people.
Crop
Opt. Temp. Vegetative
Opt. Temp. Flowering
Failure Temp.Flowering
Maize
28-35oC
18-22oC
35oC
Soybean
25-37oC
22-24oC
39oC
Wheat
20-30oC
15oC
34oC
Rice
28-35oC
23-26oC
36oC
Sorghum
26-34oC
25oC
35oC
Cotton
34oC
25-26oC
35oC
Peanut
31-35oC
20-26oC
39oC
Data are adapted from Hatfield et al., 2011.
Doubtful you’ve seen this climate threat to the global food supply on TV or a streaming service. I caught a glimpse once of temperature and agriculture on a CNN newscast, but with the “expert” calmly stating that we would just have to grow our corn in Canada, ha-ha. (Somehow, at least for rice, it’s hard to imagine India, one of the world’s largest rice producers, moving its rice production northward to the Himalaya’s, but I digress.)
Food is fundamental. If production, especially that of a global staple like rice, is impacted by rising temperatures there will be consequences. Rising prices, reduced availability, and wide-spread hunger.
But there is more to consider. Given the global dependence on rice, any change in its nutritional quality will also have effects, especially on poorer countries that rely heavily on rice as a major food source. And here we need to delve a little deeper into another aspect of climate weirding that doesn’t make it to the popular media—that rising carbon dioxide (CO2), the primary greenhouse gas, can also directly influence plant nutrition. The reasons are complicated, but in simple terms all living things consist of elements, carbon, nitrogen, phosphorous, sulfur, copper, etc., etc. A plant gets it’s carbon from the air (CO2), but everything else (nitrogen, potassium) from the soil.
And there is an imbalance. In the last 50 years, atmospheric CO2 has increased by about 30%, and is projected to increase another 50% by the end of the century. With more CO2, plants are becoming carbon rich, but nutrient poor. Nutrient poor, because while CO2 has increased in the air, nutrients in the soil have not kept pace. A perverse carb loading at the plant level.
As a consequence, rice, and many other plants, are shifting their chemistry. For example, there is a general decline in protein, in part because protein requires nitrogen. There are similar ubiquitous declines in iron and zinc, important micro-nutrients needed for human development.
Such nutritional degradation is of obvious global importance, and does, on occasion, show up on basic media when warming / weirding is mentioned, but you’d be hard pressed to find it.
Let’s move our light to another hidden bit of science. How plants can influence the air we breathe.
As humans, we like to trade things. And a large percentage of what we trade are living organisms, from fish to trees. But what began as local, regionalized trading has grown with the global population and the needs of that population—a population of 1.6 billion at the beginning of the 20th century is now ~8 billion at the beginning of the 21st. And we haven’t stopped trading. Biological trade is not inherently bad, but it represents a historically unprecedented global movement of DNA across continents, across countries, regions, towns, cities and ecosystems. And some of the DNA, when introduced, can do great harm to the environment, the economy and to human health. That harm has a name, “Invasive Species”.
Let us focus on one such plant species introduced to Eastern Europe, one that almost every American has personal experience (ACHOO!) come fall. The species is common ragweed. An invasive plant whose introduction and spread in Eastern Europe—introduced accidently through imported seeds or contaminated hay – has resulted in enormous environmental and economic losses in agriculture and public health in recent decades. In Hungary, the most important ambient biological air pollutant is: ragweed.
collecting ragweed pollen under different climates (Author’s photo)
The photo is from studies that I led looking at how ragweed pollen would respond to temperature and carbon dioxide. (If you’re curious, ragweed likes both.) Warmer temperatures, earlier Springs, later Autumns can extend its pollen season; not only extend, but increase the amount of pollen being generated. There is even some data suggesting that rising CO2 can alter pollen chemistry, making it more allergenic (REFS). Sadly, ragweed pollen doesn’t appear as temperature sensitive as that of rice, or other agricultural plants.
I wish I could say that ragweed was the exception among allergenic plants, but it’s the rule. Parthenium weed is a highly invasive species that has spread to more than 40 countries around the world. Like ragweed its pollen are highly allergenic, but it can also produce severe rashes, like poison ivy, and is known to be poisonous to livestock. It is highly aggressive, and arriving in a new location (where it has no natural enemies) can dominate landscapes, reducing biodiversity. And as with ragweed, high temperatures, longer growing seasons, heatwaves and droughts are expanding its range, and for that matter, make controlling its spread more difficult.
Such responses among invasive species will have direct impacts on air quality, especially among those (myself included) who suffer from seasonal allergies. Gasping for air is never fun.
Estimates are that pollen and seasonal asthma affects more than 24 million of us, including 6 million kids. And yet, when watching news reports of climate change, how many times have you seen a report on pollen and air quality? On increasing allergies or asthma? Once? Twice?
I could go on, (and if you need more examples, read “Greenhouse Planet”, my latest book). But my point is this: Not all of the consequences of rising carbon dioxide and climate change, warming, weirding, whatever, make for “good” TV. There is so much more to explore. So, do yourself a favor. Take a deeper dive, find out what is happening behind the scenes.
Because if we are going to rise to the challenge, we need to know what we are fighting against. Right now, the media is exemplary on showing some things, but silent on much else of importance. Watching news coverage of climate change is a bystander watching a cataclysm, and thinking, “Boy, glad I’m not experiencing THAT!”. Yet in the simplest and most basic of terms, you are, or will be, affected– from food choices to nutrition, even your allergies. And so much more.
It isn’t just about polar bears. It’s about you. Read, Understand, Act.
In a new paper, “Forest Resources of the World: Present Status and Future Prospects,” Singh et al. affirm the importance of forests for terrestrial biodiversity, provision of multiple ecosystem services, and supporting the economic well-being of approximately 1.6 billion people directly. This equals about a quarter of Earth’s population. The authors conclude that achieving global Sustainable Development Goals (SDGs), including poverty reduction, food security, and mitigating and adapting to climate change — all depend on sustaining forests.
According to the 2020 Global Forest Resource Assessment, Earth’s forested area comprises ~4.06 billion hectares, or 31% of the total land surface.More than half (54%) of all global forest area is found in five countries: the Russian Federation, Brazil, Canada, the United States, and China. Tropical forests constitute 45% of this total; boreal forests, 27%; temperate forests, 16%; and subtropical forests, 11%. An estimated 93% (3.75 billion ha) regenerate through natural processes; 7% (290 million ha) is planted forest.
The extent of global forest area has been declining for decades but the rate of loss slowed significantly between 1990 and 2020. This reflects decreased deforestation in some countries and an increase in forest area in others. The latter is due to both afforestation and also natural forest growth. However, conversion of tropical forests to agriculture continues apace. From 2010 to 2020, the net loss of forest area was highest in Africa (3.9 million ha) and South America (2.6 million ha). Increases in net forest area occurred in Asia, Oceania and Europe. The status of the top 10 countries or territories in global forest resources as of 2020 is given in Table 1.2 of the chapter. [News sources document that rapid deforestation continues in Brazil, at least.]
Several trends are concerning to those of us who value primary or undisturbed forests. First, the area of naturally regenerating forest has decreased, while the area of planted forest has expanded – but only by 123 million ha. In the last decade, the rate of increase in the area of planted forests has also slowed.
Second, total carbon stock in forests declined from 668 gigatons to 662 gt in 1990–2020. This is only 6%, but it is trending in the wrong direction. As we know, forest conservation counters climate change in two ways: conserved forests are a carbon sink, while degraded or destroyed forests are a significant source of atmospheric CO2. In fact, forests are the 2nd largest storehouses of carbon, after oceans. Global forests sequester about one-third of total CO2 emission from the combustion of fossil fuels. Almost all forest carbon is found in living biomass (44%) and soil organic matter (45%).
Costa Rican rainforest; photo by eflon via Flickr
Third, primary forests are already severely reduced and continue to shrink. Primary forests are those composed of native species, and supporting relatively undisturbed ecological processes. They are irreplaceable for sustaining biological diversity. These forests are already severely reduced – they cover only ~ 1 billion ha. Since 1990, the extent of primary forest has decreased by 81 million ha. More than half are in Brazil, Canada, and Russia.
Singh et al. report that only about 10% of the world’s forests are set aside for biodiversity conservation. Again, trends are in the wrong direction. The rate of increase in the area of forest designated largely for biodiversity conservation has slowed. On the other hand, forest areas designated for other non-extractive purposes have increased: soil and water conservation, recreation, tourism, education, research, and the protection of cultural and spiritual sites.
Singh et al. are cheered by the fact that more than 2 billion hectares are under management with well-defined management plans. The extent of forests under management plans has increased by 233 million ha since 2000.
Singh et al. say that continuously increasing anthropogenic pressure is the main cause of deforestation and forest degradation in unmanaged forests. Citing projections that the world’s population will reach almost 10 billion by 2050, they say this growth will make reconciling the need for forest conservation with the basic requirements of humans for food, shelter, and fuel more difficult than ever.
I appreciate this honesty. Too many experts interviewed on the day that the global population was estimated at 8 billion made optimistic statements about the consequences. They mentioned Earth’s carrying capacity only in reference to First World people demanding excessive resources. There was minimal discussion of humanity’s carbon footprint and no reference to ever-increasing threats to biological diversity. Nor to the fact that people in developing countries want to raise their standards of living – which entails higher demand for resources, including energy. For an example, see The Washington Post editorial, here.
On the other hand, Ruby Mellen in the Post on 15 November mentioned that, according to the World Wildlife Fund, 75% of Earth’s ice-free land has been significantly altered by people, and two-thirds of mammal, fish, reptile, and amphibian species have become endangered in the last ~50 years. Unfortunately, the on-line version of the paper doesn’t have this specific article!
fires in Siberian forest in 2016; European Space Agency
Threats to Forests: Fire
Singh et al. rank fire as the most disastrous threat, affecting biodiversity and carbon sequestration potential. According to the U.N. Food and Agriculture Organization, about 29% of the total geographical area in the world was affected by forest fires during 2001–2018; more than two-thirds of these fires occurred in Africa. U.S. media, however, focused on fires in the Amazon, temperate areas (U.S., Europe), and, sometimes, boreal forests or Australia. Singh et al. say that areas that are frequently affected by fire are prone to other types of disturbances like drought and outbreaks of insect pests.
tanoaks killed by Phytophthora ramorum in Oregon; photo by Oregon Department of Forestry
Threats to Forests: Diseases and Pests
I am glad that Singh et al. recognize the damage to forest productivity caused by disease and pest infestations. In doing so, they cite familiar sources – Clive Brasier, Peter Vitousek, Juliann Aukema, Gary Lovett, Sandy Liebhold, Kerry Britton, Bitty Roy, Hanno Seebens – regarding surges in pest attacks; the growing diversity of damaging pests; resulting changes in forest species composition and structure that impede ecosystem functions and productivity. Singh et al. follow these sources in calling for improved hygiene in nurseries, adoption of scientific silvicultural practices reducing physical damage to the vegetation, selection of genotypes that are resistant, and reinforcing national and international policies on quarantine and biosecurity measures to minimize pest impacts in the future. They also mention adoption of remote sensing technologies to detect the trees under stress and use of sentinel plantings. They list the 10 most important international agreements dealing with invasive species issues as the International Plant Protection Convention, Ramsar Convention, Convention on International Trade in Endangered Species of Wild Fauna and Flora, Convention on Migratory Species, Convention on Biological Diversity and its Cartagena Protocol on Biosafety, IUCN Invasive Species Specialist Group, World Trade Organization Agreement on Sanitary and Phytosanitary Measures, Global Invasive Species Program, and International Civil Aviation Organization, and Cartagena.
slash and burn agriculture in Bolivia; photo Neil Palmer
Threats to forests: Development Projects
Singh et al. consider development projects to be the third threat to forest conservation. Their roads, powerlines, and other linear developments cause habitat loss and fragment landscapes. In their view, environmental impact assessments and other similar requirements are not yet sufficient to safeguard sustainable use of forest resources.
Policy Responses
Singh et al. call for more inclusive forest management structures to respond to the threat climate change poses to forests, industries, and forest-dependent communities. They all for partnerships that bring together researchers from several disciplines with forest managers and local stakeholders. Geoffrey M. Williams and others (including me) advocate for similar conservation approaches. (See pre-print here.)
In this context, Singh et al. mention several reports, plans, and agreements aimed at global forest conservation. Participants in global fora have recognized the importance of forests in contributing to food security and sustainable development. Among agreements mentioned are the UN’s Strategic Plan for Forests 2030 and recommendations of the International Institute for Sustainable Development (IISD) published in 1994. The former tries to generate greater coherence, collaboration, and synergy across UN programs aimed at encouraging volunteer forest conservation by countries, international, regional, and local organizations, partners, and stakeholders. Unfortunately, they do not discuss the extent to which the 30-year old IISD recommendations have – or have not – been implemented.
They also describe Forest Landscape Restoration as an effective strategy to restore the functionality of forests.Again, the focus is on a collaborative approach aimed at integrating efforts by all forestry-related stakeholders, e.g., scientific and academic organizations, local communities, indigenous peoples, and private sectors, including forest-based enterprises and NGOs.
Also praised is rising attention to trees outside forest. This includes fostering use of trees in agroforestry systems ranging from home gardens to farm forestry systems, shelterbelts, and woodlots. This approach helps to sustain the livelihoods of rural communities and maintain a stable and secure food supply. Meanwhile, it reduces dependence on natural forests
Singh et al. say community forest management and decentralized governance have gained acceptance. They describe examples from Gambia and Rwanda. They concede that such decentralization has its own risks and challenges. For example, e the most marginalized sections of the community must be ensured adequate capacity for robust conflict resolution.
Singh et al. advocate that all nations seek to increase their forest cover; affluent countries that are hampered by physical and climatic conditions should aid poorer nations in increasing and upgrading their forest cover. They suggest “recognition” and encouragement of countries that maintain forest cover above 30% of territory.
Singh, M., N.N. Shahina, S. Das, A. Arshad, S. Siril, D. Barman, U. Mog, P. Panwar, G. Shukla, and S. Chakravarty. 2022. Forest Resources of the World: Present Status and Future Prospects. In Panwar, P., G. Shukla, J.A. Bhat, S. Chakravarty. 2022. Editors. Land Degradation Neutrality: Achieving SDG 15 by Forest Management; ISBN 978-981-19-5477-1 ISBN 978-981-19-5478-8 (eBook)
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
TACF back-crossed chestnut growing in southern Fairfax County, Virginia
same chestnut, dying in 2021; probably from Phytopthora root rot
In the first half of the 20th Century, American chestnut (Castanea dentata) was functionally extirpated from US forests east of the Mississippi River by chestnut blight, caused by a fungus from Asia, Cryphonectria parasitica. Today, only 10% of the pre-blight chestnut population remains, most as root sprouts less than 2.5 cm dbh (Dalgleish et al. 2015; full citation at the end of the blog).
Volunteer organizations — with recent help from federal and state agencies – have worked for more than a century to develop chestnut trees resistant to the blight. Their aim is to restore the species to the forest. Their decades of hybridization efforts now appear unlikely to produce a highly blight-resistant chestnut with a genome that is predominantly American, so TACF now plans to incorporate the use of transgenic techniques to enhance resistance to the blight fungus.
However, restoration of chestnut requires addressing a second Asian pathogen: Phytophthora cinnamomi, which causes a fatal root disease. Several studies indicate that up to 80% of seedlings are killed. The pathogen is widespread in soils south of 40o North Latitude, which falls just north of the Maryland-Pennsylvania line. Thus, P. cinnamomi occupies the southern half of American chestnut’s former range. Scientists expect this pathogen to move north in response to the warming climate; indeed, some project that the root disease could reach throughout the entire current chestnut rangeby 2080.
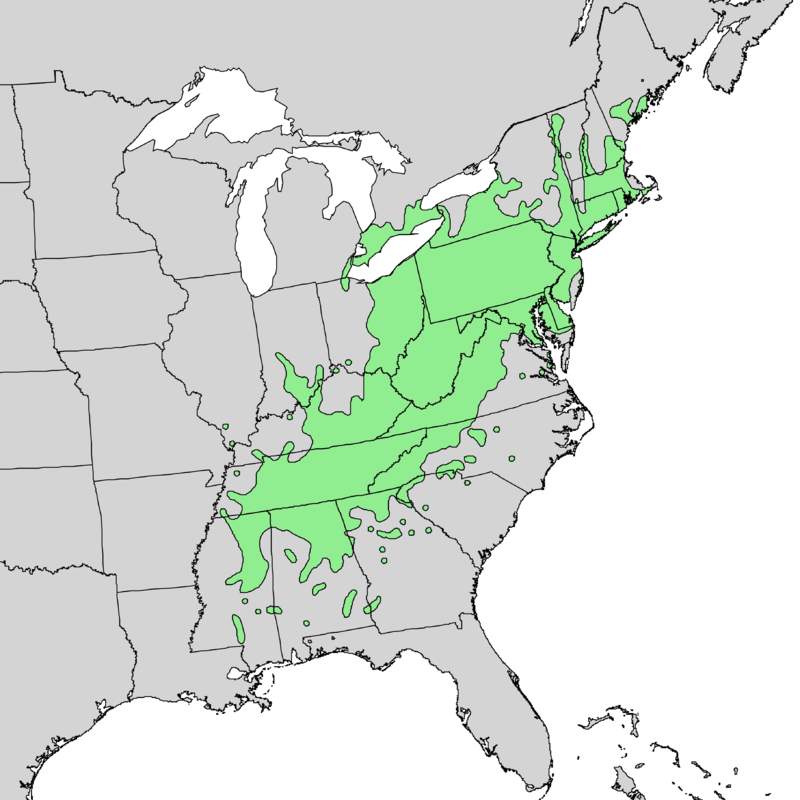
historic range of American chestnut
Gustafson et al. 2022 modelled chestnut’s vulnerability to P. cinnamomi to current and expected environmental conditions in two state forests in the Appalachians of western Maryland to evaluate the probable impact of the root disease on efforts to restore the tree species.
They found that root rot greatly reduced chestnut biomass on the landscape, even when resistance to root rot was at the target level for selection of root rot-resistant chestnut families using traditional breeding methods.
Gustafson et al. 2022 recommend that chestnut restoration apply the following strategies:
Locate restoration plantings at latitudes, elevations, and sites where root rot is not expected to be present well into the future. This probably means sites in the Northeastern US and Canada (Burgess et al. 2017)
Enhance the planting stock’s resistance to P. cinnamomi through breeding.
Identify soil conditions, including soil microbes, that suppress the pathogen or protect tree roots.
Since planting stock – both bareroot and containerized – can transmit P. cinnamomi, either raise seedlings in nurseries located outside the pathogen’s current range or rely on direct seeding. These strategies have their own downsides. Restricting locations of nurseries might complicate efforts to ensure seedlings are adapted to local conditions in the restoration area and seeds would need to be protected from seed predators.
The authors specify these additional important conditions:
Planting locations: while Canada is currently outside the range of American chestnut, the same climatic warming that will facilitate northward spread of P. cinnamomi will probably allow the tree to thrive farther north (Barnes and Delborne 2019). Perhaps the tree’s range will shift farther north than the pathogen’s.
Breeding: some resistance to Phytophthora root rot has been found in families providing blight resistance used in The American Chestnut Foundation (TACF) breeding program. TACF now plans to cross individuals from those families with transgenic blight-resistant chestnut to combine both resistances.
Soils: P. cinnamomi is favored by compacted soils with poor aeration or that tend to remain saturated. These include heavy clay soils and those highly disturbed by agriculture or mining. Restoration sites should be non-disturbed, well-drained sites. (This recommendation contradicts others’ proposals that chestnuts be planted on reclaimed mining sites.) Silvicultural management should also minimize environmental stresses.
that dying chestnut trying to reproduce!
Restoring chestnut will be challenging in any case: successful restoration requires chestnut trees that can compete successfully in the forest and adapt to conditions which are now quite different from those a century ago when the species was dominant. These include abiotic factors, e.g., climate and atmospheric CO2 levels; and biotic factors, e.g., different forest pests and invasive plant species.
In an earlier publication, Gustafson and colleagues (Gustafson et al. 2018) modelled the effects of warmer temperature and elevated atmospheric CO2 levels on chestnut’s growth and competition and the tree’s adaptation to natural and anthropogenic disturbances. They concluded that aggressive restoration programs – involving clearcutting, then planting chestnuts – could restore chestnut as an important component of forested ecosystems in the Appalachian Mountains.
However, this earlier study did not consider the effects of Phytophthora root rot. The 2022 study demonstrates that these recommendations are probably applicable only to the northernmost portion of former chestnut range, outside the areas infested by Phytophthora root rot, unless breeding is successful in substantially increasing resistance to root rot.
Several studies indicate American chestnut is highly susceptible to P. cinnamomi; rates of root rot induce mortality of 80% or higher have been documented. TACF has found that hybrid chestnut families selected for root rot resistance have a mortality rate of about 45%. Even with this level of tolerance, the model shows that chestnut could not regain anything approaching its former abundance on the landscape. Since the threat of P. cinnamomi to chestnut restoration has become evident, TACF is assessing how to integrate increased tolerance to root rot into their larger blight resistance breeding program (Westbrook et al. 2019).
Soil properties – texture, land use, drainage, waterlogging, drought, temperature, and water-holding capacity – influence infection. So does weather: a single heavy rain event might saturate soil sufficiently to facilitate a P. cinnamomi infection. For these reasons, climate change is expected to exacerbate its geographic spread and pathogenicity.
The sites used in both studies are at the center of chestnut’s former range, which is also at the northern edge of the root rot pathogen’s range. However, the two sites differ in important ways, especially in rainfall and soils. The researchers considered one a mesic site and the other, xeric.
Their 2022 model showed that root rot caused a dramatic reduction in chestnut biomass on both the mesic and xeric sites. Apparently temperature and wetness levels offset each other. That is, higher soil temperatures intensified P. cinnamomi virulence at the xeric site sufficiently to overcome its relative soil dryness. At the mesic site, soil temperature sometimes dropped to levels that are lethal to Phytophthora. On the whole, then, climate change is expected to intensify P. cinnamomi infection rates on both sites and reduce the number of sites where the pathogen is absent.
Gustafson et al. (2022) discuss several assumptions and data gaps that require further study.
SOURCES
Barnes, J.C. and Delborne, J.A., 2019. Rethinking restoration targets for American chestnut using species distribution modeling. Biodiversity and Conservation, 28(12), pp.3199-3220.
Burgess, T.I., Scott, J.K., Mcdougall, K.L., Stukely, M.J., Crane, C., Dunstan, W.A., Brigg, F., Andjic, V., White, D., Rudman, T. and Arentz, F., 2017. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Global Change Biology, 23(4), pp.1661-1674.
Dalgleish, H.J., Nelson, C.D., Scrivani, J.A. and Jacobs, D.F., 2015. Consequences of shifts in abundance and distribution of American chestnut for restoration of a foundation forest tree. Forests, 7(1), p.4.
Gustafson, E.J., B.R. Miranda, T.J. Dreaden, C.C. Pinchot, D.F. Jacobs. 2022. Beyond blight: Phytophthora root rot under climate change limits populations of reintro Am chestnut Ecosphere. 2022;13:e3917.
Gustafson, E.J., A.M.G. De Bruijn, N. Lichti, D.F. Jacobs, B.R. Sturtevant, D.M. Kashian, B.R. Miranda, and P.A. Townsend. 2018. “Forecasting Effects of Tree Species Reintroduction Strategies on Carbon Stocks in a Future without Historical Analog.” Global Change Biology 24: 5500–17. https://doi.org/10.1111/gcb.14397
Westbrook, Jared W., et al. “Resistance to Phytophthora cinnamomi in American chestnut (Castanea dentata) backcross populations that descended from two Chinese chestnut (Castanea mollissima) sources of resistance.” Plant disease 103.7 (2019): 1631-1641.
Posted by Faith Campbell
[An earlier version of this blog has now been corrected, with additional sources added. I think Cornelia Pinchot, USFS, for the corrections.]
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Horton House on Jekyll Island, Georgia before laurel wilt killed the giant redbay trees; photo by F.T. Campbell
Close to four hundred tree species native to the United States are at risk of extinction. The threats come mainly from non-native insects and diseases – a threat we know gets far too little funding, policy attention, and research.
As Murphy Westwood, Vice President of Science and Conservation at the Morton Arboretum, which led the U.S. portion of a major new study, said to Gabriel Popkin, writing for Science: “We have the technology and resources to shift the needle,” she says. “We can make a difference. We have to try.”
Staggering Numbers
More than 100 tree species native to the “lower 48” states are endangered (Carrero et al. 2022; full citation at the end of this blog). These data come from a global effort to evaluate tree species’ conservation status around the world. I reported on the global project and its U.S. component in September 2021. This month Christina Carrero and colleagues (full citation at the end of this blog) published a summary of the overall picture for the 881 “tree” species (including palms and some cacti and yuccas) native to the contiguous U.S. (the “lower 48”).
This study did not address tree species in Hawai`i or the U.S. Pacific and Caribbean territories. However, we know that another 241 Hawaiian tree species are imperiled (Megan Barstow, cited here).
Assessing Threats: IUCN, NatureServe, and CAPTURE
Carrero and colleagues assessed trees’ status by applying methods developed by IUCN and NatureServe. (See the article for descriptions of these methods.) These two systems consider all types of threats. Meanwhile, three years ago Forest Service scientists assessed the specific impacts of non-native insects and pathogens on tree species in the “lower 48” states and Alaska in “Project CAPTURE” (Conservation Assessment and Prioritization of Forest Trees Under Risk of Extirpation). All three systems propose priorities for conservation efforts. For CAPTURE’s, go here.
Analyses carried out under all three systems (IUCN, NatureServe, and CAPTURE) concur that large numbers of tree species are imperiled. Both IUCN and CAPTURE agree that non-native insects and pathogens are a major cause of that endangerment. While the overall number of threatened species remained about the same for all three systems, NatureServe rated threats much lower for many of the tree species that IUCN and CAPTURE considered most imperiled.
This difference arises from the criteria used to rate a species as at risk. IUCN’s Criterion A is reduction in population size. Under this criterion, even extremely widespread and abundant species can qualify as threatened if the population declines by at least 30% over three generations in the past, present, and/or projected future. NatureServe’s assessment takes into account rapid population decline, but also considers other factors, for example, range size, number of occurrences, and total population size. As a result, widespread taxa are less likely to be placed in “at risk” categories in NatureServe’s system.
In my view, the IUCN criteria better reflect our experience with expanding threats from introduced pests. Chestnut blight, white pine blister rust, dogwood anthracnose, emerald ash borer, laurel wilt disease, beech leaf disease, and other examples all show how rapidly introduced pathogens and insects can spread throughout their hosts’ ranges. (All these pests are profiled here . ) They can change a species’ conservation status within decades whether that host is widespread or not.
Which Species Are at Risk: IUCN
Carrero and colleagues found that under both IUCN and NatureServe criteria, 11% to 16% of the 881 species native to the “lower 48” states are endangered. Another five species are possibly extinct in the wild. Four of the extinct species are hawthorns (Crataegus); the fifth is the Franklin tree (Franklinia alatamaha) from Georgia. A single specimen of a sixth species, an oak native to Texas (Quercus tardifolia),was recently re-discovered in Big Bend National Park.
Franklinia (with Bachman’s warbler); both are extinct in the wild; painting by John Jacob Audubon
The oak and hawthorn genera each has more than 80 species. Relying on the IUCN process, Carrero and colleagues found that a significant number of these are at risk: 17 oaks (20% of all species in the genus); 29 hawthorns (34.5% percent). A similar proportion of species in the fir (Abies), birch (Betula), and walnut (Juglans) genera are also threatened.
Other genera have an even higher proportion of their species under threat, per the IUCN process:
all species in five tree genera, including Persea (redbay, swampbay) and Torreya (yews);
two-thirds of chestnuts and chinkapins (Castanea), and cypress (Cupressus);
almost half (46.7%) of ash trees (Fraxinus).
Pines are less threatened as a group, with 15% of species under threat. However, some of these pines are keystone species in their ecosystems, for example the whitebark pine of high western mountains.
Carrero et al. conclude that the principal threats to these tree species are problematic and invasive species; climate change and severe weather; modifications of natural systems; and overharvest (especially logging). Non-native insects and pathogens threaten about 40 species already ranked by the IUCN criteria as being at risk and another 100 species that are not so ranked. Climate change is threatening about 90 species overall.
range of black ash
Considering the invasive species threat, Carrero and colleagues cite specifically ash trees and the bays (Persea spp.). In only 30 years, the emerald ash borer has put five of 14 ash species at risk. All these species are widespread, so they are unlikely to be threatened by other, more localized, causes. In about 20 years, laurel wilt disease threatens to cause extinction of all U.S. tree species in the Persea genus.
Carrero and colleagues note that conservation and restoration of a country’s trees and native forests are extremely important in achieving other conservation goals, including mitigating climate change, regulating water cycles, removing pollutants from the air, and supporting human well-being. They note also forests’ economic importance.
As I noted above, USFS scientists’ “Project CAPTURE” also identified species that deserve immediate conservation efforts.
Where Risk Assessments Diverge
All three systems for assessing risks agree about the severe threat to narrowly endemic Florida torreya and Carolina hemlock.
With three risk ranking systems, all can agree (as above), all can disagree, or pairs can agree in four different ways. Groups of trees fall into each pair, with various degrees of divergence. Generally, only two of the three systems agree on more widespread species:
black ash: IUCN and Project CAPTURE prioritize this species. NatureServe ranked it as “secure” (G5) as recently as 2016.
whitebark pine: considered endangered by IUCN, “vulnerable” (G3) by NatureServe. The US Fish and Wildlife Service has proposed listing the species as “threatened” under the Endangered Species Act. https://www.fws.gov/species-publication-action/endangered-and-threatened-wildlife-and-plants-threatened-species-18 However, Project CAPTURE does not include it among its highest priorities for conservation. Perhaps this is because there are significant resistance breeding and restoration projects already under way.
tanoak: considered secure by both IUCN and NatureServe, but prioritized by Project CAPTURE for protection.
dead tanoak in Curry County, Oregon; photo by Oregon Department of Forestry
Carrero notes the divergence between IUCN and NatureServe regarding ashes. Four species ranked “apparently secure” (G4) by NatureServe (Carolina, pumpkin, white, and green ash) are all considered vulnerable by IUCN. They are also prioritized by Project CAPTURE. I have described the impact of the emerald ash borer on black ash. Deborah McCullough, noted expert on ash status after invasion by the emerald ash borer, also objects to designating this species as “secure” (pers. comm.).
Port-Orford cedar is currently ranked as at risk by IUCN and Project CAPTURE, but not NatureServe. Growing success of the restoration breeding project has prompted IUCN to change the species’ rank from “vulnerable” to “near threatened”. IUCN is expected to reclassify it as of “least concern” in about a decade if breeding efforts continue to be successful (Sniezko presentation to POC restoration webinar February 2022).
While these differing detailed assessments are puzzling, the main points are clear: several hundred of America’s tree species (including many in Hawai`i, which – after all – is our 50th state!) are endangered and current conservation and restoration efforts are inadequate.
Furthermore, a tree species loses its function in the ecosystem long before it becomes extinct. It might still be quite numerous throughout its range – but if each individual has shrunken in size it cannot provide the same ecosystem services. Think of thickets of beech root sprouts – they cannot provide the bounteous nut crops and nesting cavities so important to wildlife. Extinction is the extreme. We should act to conserve species much earlier.
YOU CAN HELP!
Congress is considering the next Farm Bill – which is due to be adopted in 2023. Despite its title, this legislation has often provided authorization and funding for forest conservation (for example, the US Forest Service’ Landscape Scale Restoration Program).
There is already a bill in the House of Representatives aimed at improving the US Department of Agriculture’s prevention and early detection/rapid response programs for invasive pests. Also, it would greatly enhance efforts to restore decimated tree species via resistance breeding, biocontrol, and other strategies. This bill is H.R. 1389.
The bill was introduced by Rep. Peter Welch of Vermont, who has been a solid ally and led on this issue for several years. As of August 2022, the bill has seven cosponsors, most from the Northeast: Rep. Mike Thompson [CA], Rep. Chellie Pingree [ME], Reps. Ann M. Kuster and Chris Pappas [NH], Rep. Elise Stefanik [NY], Rep. Deborah K. Ross [NC], Rep. Brian Fitzpatrick [PA].
Please write your Representative and Senators. Urge them to seek incorporation of H.R. 1389 in the 2023 Farm Bill. Also, ask them to become co-sponsors for the House or Senate bills. (Members of the key House and Senate Committees are listed below, along with supporting organizations and other details.)
Details of the Proposed Legislation
The Invasive Species Prevention and Forest Restoration Act [H.R. 1389]
Expands USDA APHIS’ access to emergency funding to combat invasive species when existing federal funds are insufficient and broadens the range of actives that these funds can support.
Establishes a grant program to support research on resistance breeding, biocontrol, and other methods to counter tree-killing introduced insects and pathogens.
Establishes a second grant program to support application of promising research findings from the first grant program, that is, entities that will grow large numbers of pest-resistant propagules, plant them in forests – and care for them so they survive and thrive.
[A successful restoration program requires both early-stage research to identify strategies and other scientists and institutions who can apply that learning; see how the fit together here.]
Mandates a study to identify actions needed to overcome the lack of centralization and prioritization of non-native insect and pathogen research and response within the federal government, and develop national strategies for saving tree species.
Incorporating the provisions of H.R. 1389 into the 2023 Farm Bill would boost USDA’s efforts to counter bioinvasion. As Carrera and colleagues and the Morton Arboretum study on which their paper is based demonstrate, our tree species desperately need stronger policies and more generous funding. Federal and state measures to prevent more non-native pathogen and insect pest introductions – and the funding to support this work – have been insufficient for years. New tree-killing pests continue to enter the country and make that deficit larger –see beech leaf disease here. Those here, spread – see emerald ash borer to Oregon.
For example, funding for the USDA Forest Service Forest Health Protection program has been cut by about 50%; funding for USFS Research projects that target 10 high-profile non-native pests has been cut by about 70%.
H.R. 1389 is endorsed by several organizations in the Northeast: Audubon Vermont, the Maine Woodland Owners Association, Massachusetts Forest Alliance, The Nature Conservancy Vermont, the New Hampshire Timberland Owners Association, Vermont Woodlands Association, and the Pennsylvania Forestry Association.
Also, major forest-related national organizations support the bill: The American Chestnut Foundation (TACF), American Forest Foundation, The Association of Consulting Foresters (ACF), Center for Invasive Species Prevention, Ecological Society of America, Entomological Society of America, National Alliance of Forest Owners (NAFO), National Association of State Foresters (NASF), National Woodland Owners Association (NWOA), North American Invasive Species Management Association (NAISMA), Reduce Risk from Invasive Species Coalition, The Society of American Foresters (SAF).
HOUSE AND SENATE AGRICULTURE COMMITTEE MEMBERS – BY STATE
STATE
Member, House Committee
Member, Senate Committee
Key members * committee leadership # forestry subcommittee leadership @ cosponsor of H.R. 1389
Alabama
Barry Moore
Arizona
Tom O’Halleran
Arkansas
Rick Crawford
John Boozman*
California
Jim Costa Salud Carbajal Ro Khanna Lou Correa Josh Harder Jimmie Panetta Doug LaMalfa
Colorado
Michael Bennet #
Connecticut
Jahana Hayes
Florida
Al Lawson Kat Cammack
Georgia
David Scott * Sanford Bishop Austin Scott Rick Allen
Raphael Warnock Tommy Tuberville
Illinois
Bobby Rush Cheri Bustos Rodney Davis Mary Miller
Richard Durbin
Note that the report was led by scientists at the Morton Arboretum – in Illinois!
Indiana
Jim Baird
Mike Braun
Iowa
Cindy Axne Randy Feenstra
Joni Ernst Charles Grassley
Kansas
Sharice Davids Tracey Mann
Roger Marshall#
Kentucky
Mitch McConnell
Maine
Chellie Pingree @
Massachusetts
Jim McGovern
Michigan
Debbie Stabenow *
Minnesota
Angie Craig Michelle Fischbach
Amy Klobuchar Tina Smith
Mississippi
Trent Kelly
Cindy Hyde-Smith
Missouri
Vicky Hartzler
Nebraska
Don Bacon
Deb Fischer
New Hampshire
Ann McLane Kuster @
New Jersey
Cory Booker
New Mexico
Ben Ray Lujan
New York
Sean Patrick Maloney Chris Jacobs
Kristen Gillibrand
North Carolina
Alma Adams David Rouzer
North Dakota
John Hoeven
Ohio
Shontel Brown Marcy Kaptur Troy Balderson
Sherrod Brown
Pennsylvania
Glenn Thompson
South Dakota
Dusty Johnson
John Thune
Tennessee
Scott DesJarlais
Texas
Michael Cloud Mayra Flores
Vermont
Patrick Leahy
Virginia
Abigail Spanberger #
Washington
Kim Schreir
SOURCES
Christina Carrero, et al. Data sharing for conservation: A standardized checklist of US native tree species and threat assessments to prioritize and coordinate action. Plants People Planet. 2022;1–17. wileyonlinelibrary.com/journal/ppp3
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Pinus radiata plantation in New Zealand; photo by Jon Sullivan
As countries and conservation organizations ramp up tree planting as one solution to climate change, I worry that many of the plantings will use species not native to the region – with the risk of promoting more bioinvasions. My second fear is that inadequate attention will be paid to ensuring that the propagules thrive.
Warning from New Zealand
New Zealand has adopted a major afforestation initiative (“One Billion Trees”). This program is ostensibly governed by a policy of “right tree, right place, right purpose”. However, Bellingham et al. (2022) [full citation at end of blog] say the program will probably increase the already extensive area of radiata pine plantations and thus the likelihood of exacerbated invasion. They say the species’ potential invasiveness and its effects in natural ecosystems have not been considered.
Bellingham et al. set out to raise the alarm by evaluating the current status of radiata, or Monterrey, pine (Pinus radiata) in the country. They note that the species already occupies ~1.6 M ha; the species makes up 90% of the country’s planted forests. Despite the species having been detected as spreading outside plantations in 1904, it is generally thought not to have invaded widely.
The authors contend that, to the contrary, radiata pine has already invaded several grasslands and shrublands, including three classes of ecosystems that are naturally uncommon. These are geothermal ecosystems, gumlands (infertile soils that formerly supported forests dominated by the endemic and threatened kauri tree Agathis australis), and inland cliffs. Invasions by pines – including radiata pine – are also affecting primary succession on volcanic substrates, landslides on New Zealand’s steep, erosion-prone terrain, and coastal sand dunes. Finally, pine invasions are overtopping native Myrtaceae shrubs during secondary succession. Bellingham et al. describe the situation as a pervasive and ongoing invasion resulting primarily from spread from plantations to relatively nearby areas.
kauri; photo by Natalia Volna, iTravelNZ
The New Zealanders cite data from South America and South Africa on the damaging effects of invasions by various pine species, especially with respect to fire regimes.
Furthermore, their modelling indicates that up to 76% of New Zealand’s land area is climatically capable of supporting radiata pine — most of the country except areas above 1000 m in elevation or receiving more than 2000 mm of rainfall per year. That is, all but the center and west of the South Island. This model is based on current climate; a warmer/drier climate would probably increase the area suitable to radiata pine.
These invasions by radiata pine have probably been overlooked because the focus has been on montane grasslands (which are invaded by other species of North American conifers). [See below — surveys of knowledge of invasive plants’ impacts.]
Bellingham et al. recognize the economic importance of radiata pine. They believe that early detection of spread from plantations and rapid deployment of containment programs would be the most effective management strategy. They therefore recommend
1) taxing new plantations of non-indigenous conifers to offset the costs of managing invasions, and
2) regulating these plantations more strictly to protect vulnerable ecosystems.
They also note several areas where additional research on the species’ invasiveness, dispersal, and impacts is needed.
Survey of Awareness of Invasive Plants
A few months later a separate group of New Zealand scientists published a study examining tourists’ understanding of invasive plant impacts and willingness to support eradication programs (Lovelock et al.; full citation at end of the blog). One of the invasive plant groups included in the study are conifers introduced from North America and Europe. These conifers are invading montane grasslands, so they are not the specific topic of the earlier article. The other is a beautiful flowering plant, Russell lupine. These authors say that both plant groups have profound ecological, economic, and environmental impacts. However, the conifers and lupines are also highly visible at places valued by tourists. Lovelock et al. explored whether the plants’ familiarity – and beauty – might affect how people reacted to descriptions of their ecosystem impacts.
Visitors from elsewhere in New Zealand were more aware of invasive plants’ impacts and more willing to support eradication programs for these species specifically. Asian visitors had lower awareness and willingness to support eradication of the invasives than tourists from the United Kingdom, Europe, or North America. This pattern remained after the tourists were informed about the plants’ ecological impacts. All groups were less willing to support eradication of the attractive Russell lupine than the conifers.
Conifers invading montane grasslands are perhaps the most publicized invasive plants in New Zealand [as noted above]. Lovelock et al. report that New Zealand authorities have spent an estimated $NZ166 million to eradicate non-native conifers over large tracts of land on the South Island. Still, only about half the New Zealand visitors surveyed were aware of the ecological problems caused by wild conifers.
invasive lupines in New Zealand; photo by Michael Button via Flickr
Russell lupine (Lupinus × russellii) is invading braided river systems, modifying river flows, reducing nesting site availability for several endangered birds, and provides cover for invasive predators. While initially planted in gardens, the lupines were soon being deliberately spread along the roads to ‘beautify’ the landscape. Foreign tourists often specifically seek river valley invaded by the lupine because pictures of the floral display appear in both official tourism promotional material & tourist-related social media. It is not surprising, then, that even among New Zealanders, only a third were aware of the lupines’ environmental impacts.
The oldest participants (those over 60) had the lowest acceptance of wild conifers. Participants 50–59 years old were most aware of ecological problems caused by wild conifers. Participants 30–39 years old showed the highest acceptance of wild conifers and lowest awareness of ecological issues.
Female participants showed a higher preference for the landscape with wild conifers (45.90%) than males (36.89%). Female participants were also half as aware of ecological problems (25.62% v. 46.12% among male participants).
Nearly all survey participants (96.1%) preferred the landscape with flowering lupine; only 19.4% were aware of associated ecological problems. New Zealand domestic visitors were more aware. After the impacts of lupines were explained, half decided to support eradication. However, the same proportion of all survey participants (42.5%) still wanted to see lupines in the landscape.
Once again, participants older than 50 were more aware of ecological problems arising from lupine invasions. Both men and women greatly preferred the landscape with Russell lupins.
While the authors do not explore the ramifications of the finding that younger people are less aware of invasive species impacts, I think they bode ill for future protection of the country’s unique flora and fauna. They did note that respondents had a high level of acceptance overall for these species on the New Zealand landscapes.
While the study supported use of simple environmental messaging to influence attitudes about invasive species, also showed that need to consider such social attributes as nationality and ethnicity. So Lovelock et al. call for investigation of how and why place of origin and ethnicity are important in shaping attitudes towards invasives. Conveying conservation messages will be more difficult because tourist materials often contain photographs of the lupines. Much of this information comes from informal media such as social media, which are beyond the control of invasive species managers.
SOURCES
Bellingham, P.J., E.A. Arnst, B.D. Clarkson, T.R. Etherington, L.J. Forester, W.B. Shaw, R. Sprague, S.K. Wiser, and D.A. Peltzer. 2022. The right tree in the right place? A major economic tree species poses major ecological threats. Biol Invasions Vol.: (0123456789) https://doi.org/10.1007/s10530-022-02892-6
Lovelock B., Y. Ji, A. Carr, and C-J. Blye. 2022. Should tourists care more about invasive species? International and domestic visitors’ perceptions of invasive plants and their control in New Zealand. Biological Invasions (2022) 24:3905–3918 https://doi.org/10.1007/s10530-022-02890-8
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
loblolly pine – tree species showing highest value in this study; via Flickr
More scientists are examining the importance of American forests in providing ecosystem services – and the threat to those values raised by non-native pests and other factors. This is a broader perspective than used in the past – and it includes climatechange. also here
Jeannine Cavender-Bareu and colleagues (full citation at the end of this blog) found that changes in the abundance and composition of US trees have the potential to undermine the benefits and societal values derived from those forests now. They examined threats associated with increasing invasive pests and pathogens, greater frequency of major fires, and climate change. Together, these constitute a complex set of global change drivers – and the impact of each is accelerating.
The authors tried to measure the impact of these forces on forests’ ability to provide five key ecosystem services. Two are “regulating” services—regulation of climate and air quality. The other three are “provisioning” services—production of wood products, food crops, and Christmas trees.
Unfortunately, they could not find sufficient data to analyze five other ecosystem services, which are equally or more important. They include both regulatory and provisioning services: water management, such as erosion control, flood and storm surge regulation; urban heat island regulation and energy savings; providing habitats for species (biodiversity); recreation; or ornamental, spiritual, and aesthetic values.
Cavender-Bareu and colleagues concluded that the value of the five analyzed services provided by 400 tree species across the contiguous United States over the years 2010-2012 is $114 billion per year. The non-market “regulatory” values far exceeds their current commercial value.
Climate regulation via carbon storage in tree biomass provides 51% of this net annual value;
Human health improvements linked to trees’ filtering of air pollution provide an additional 37% of the annual net value.
Provisioning services, such as wood products, fruit and nut crops, and Christmas trees, provide only 12% of the net annual value. (By my calculation, wood products constituate almost three-quarters of this sum.)
The authors then tried to identify which tree lineages, e.g., taxonomic families, genera, or species, provide the greatest proportion of each of these ecosystem services. They also identified threats to these lineages. Together, this knowledge allows managers to target forestry management practices to the specific lineages within a landscape where ecosystem service are most at risk.
Table 1 in the article ranks 10 tree genera by the aggregate net value they provide: pine, oak, maple, Douglas-fir, hemlock, cherry/almond, spruce, hickories, yellow or tulip poplar, and ash. The table also provides separate dollar values for each of the five benefits.
Two lineages—pines and oaks — provide 42% of the value of these services (annually, pines = $25.4 billion; oaks = $22.3 billion). They note that these high values result from the large number of pine and oak species occupying diverse ecological niches. Oaks have the highest annual values for climate moderation or carbon storage ($10.7 billion) and air quality regulation ($11 billion). Oaks’ air quality regulation value reflects three factors: the genus’ abundance, the trees’ size, and the large numbers planted in cities and suburbs, that is, near human populations affected by pollution. Other than this issue of location, closely related tree species tend to have similar air quality regulation values.
Many lineages provide wood products, but the amounts vary widely among related species. Pines dominate annual net revenues from wood products at $7.4 billion, due in part to their high volume and higher than average price. The most valuable species in the context of this study’s set of ecosystem services are loblolly pine (Pinus taeda) and Douglas-fir (Pseudotsuga menziesii).
Edible fruits are concentrated in two lineages — family Rosaceae, especially genera Prunus and Malus; and family Rutaceae, genus Citrus. This category demonstrates the impact of disease: annual net returns from citrus products were actually negative during the 2010 – 2012 period due to abnormally low market prices and the prevalence of citrus greening disease in Florida, Arizona and California.
northern red oak – high value for timber & carbon sequestration; photo by dcrjsr via Wikimedia
Trees at Risk
As climate change progresses, the mix of tree species that provide critical ecosystem services will be altered—with unknown consequences. There could be increases in some services but also widely-expected losses in ecosystem benefits and human well-being.
An estimated 81% of tree species are projected to have at least 10% of their biomass exposed to climates outside their current climate envelope, impacting nearly 40% of total tree biomass in the contiguous U.S. An estimated 40% of species are projected to face increasing fire frequency. In both cases, individual species’ vulnerability depends more on where that species grows than on its genetic lineage. This analysis demonstrates a threatening interaction between these two disturbance agents: the species most valuable for carbon storage are also the most at risk from the increasing fire threat.
Known (established) pests threaten 16% of tree species and potentially affect up to 40% of total tree biomass. At greatest risk are the oak and pine genera (due to mountain pine beetle and oak wilt) plus most of the crop species. The authors cite chestnut blight and Dutch elm disease as examples of pests decimating once-dominant tree species — ones provided many services. In contrast to climate and fire risks, genetic relationships explain much of the risk of pest damage because most pests attack individual species, genera, or families. (There are exceptions – sudden oak death and the Fusarium fungi vectored by invasive shot hole borers attack species across a wide range of families.)
Cavender-Bareu and colleagues conclude that major losses to pest attack of dominant species and lineages that currently provide high-value ecosystem services would undermine forest capacity to provide important benefits—at least for decades. They note that pest threats appear to be increasing partially as a consequence of climate change, demonstrating that multiple threats can interact and exacerbate outcomes. They say policy interventions aimed at slowing pests’ spread will probably be necessary to preserve the ecosystem service of climate and air quality regulation.
The high diversity of tree taxa in U.S. forests might buffer losses of ecosystem service if the most valuable lineages (oaks and pines) are compromised. However, other species will be needed to fill the voids their loss creates. Ensuring this possibility will require intentional management of forests and trees in the face of myriad and simultaneous threats.
The authors also show how tree-provided ecosystem services are distributed across the U.S. depending largely on the locations of forests, tree plantations, and orchards. Climate and air quality regulation occurs everywhere forests grow. Timber production is concentrated in a subset of the regions that also produce high climate regulation and air pollution removal, including the Southeast, Pacific Northwest, Northeast, and Upper Midwest.
The most valuable tree crops are grown on the coasts, often where forests do not grow—e.g., California; and in several Southwestern, Southern, and Eastern states.
Cavender-Bareu and colleagues found that climate change threatens species in all parts of the continent. Wildfires are expected to increase especially in California and the Intermountain West, which they say coincides with high annual storage of carbon. (This finding is opposite from those of Quirion et al. (2021) which pointed to the slow growth of pines in this region as reducing carbon storage potential.)
Cavender-Bareu and colleagues found that pest threats are highest in the Southwest and Southeast. These pests (native and non-native) are predicted to disproportionally affect species that generate high annual net values for climate regulation, air quality regulation, and wood products – e.g., pines and oaks. As noted above, these values are driven by their abundance. They note that mountain pine beetle and oak wilt have not yet reached areas with high wood product production in Northeast and Southeast.
Other studies (see Aukema et al. 2010) and here & here show that the greatest threats from non-native pests are to the Northeast/Midwest, and the Pacific coast – and Hawai`i & here.
Rock Creek Park, Washington, D.C. – an urban forest! photo by Bonnachaven
Cavender-Bareu and colleagues’ analysis advances our understanding of the threat several change drivers pose to benefits Americans receive from our forests. However, we must remember that some of the most important ecosystem services were not included because of insufficient data. Missing services:
1) most urban ecosystems. Inclusion of urban trees in the analysis would significantly increase the value of avoided health damage due to tree-based removal of air pollution. Urban trees also help regulate climate change (Nowak et al. estimate 643 M Mg of carbon are stored in urban areas, at a value of $2.31 billion annually).
2) many other regulating ecosystem services, such as erosion control, flood regulation, storm surge regulation, urban heat island regulation, energy savings due to shade, and species habitat / biodiversity.
3) recreation, ornamental, spiritual, and aesthetic values.
A complete accounting would also require estimates of the damage trees cause and the cost of their maintenance. For example, the full cost of irrigating almond trees; allergies and irritations due to tree pollen and sap; injuries to people and property caused by falling trees and limbs; trees’ role in spreading fires; trees’ contribution to volatile organic compounds (a pollutant).
The estimated annual values of the climate and air quality regulation have large uncertainty. These arise from uncertainty re: the social cost of carbon, the value of a statistical life, and uncertainty in the air pollution dose–mortality response function. The estimated annual values of the provisioning services are more precise because they are calculated from the market price for the per unit value of tree crops, wood products, and Christmas trees, as well as reliable data on production volume.
SOURCES
Aukema, J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, & S.J. Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the Continental United States. Bioscience. December 2010 / Vol. 60 No. 11
Cavender-Bareu, J.M., E. Nelson, J.E. Meireles, J.R. Lasky, D.A. Miteva, D.J.Nowak, W.D. Pearse, M.R. Helmus, A.E. Zanne, W.F. Fagan, C. Mihiar, N.Z. Muller, N.J.B. Kraft, S. Polasky. 2022. The hidden value of trees — Quantifying the ecosystem services of tree lineages and their major threats across the contiguous. PLOS Sustainability and Transformation April 5, 2022.
Quirion BR, Domke GM, Walters BF, Lovett GM, Fargione JE, Greenwood L, Serbesoff-King K, Randall JM & Fei S (2021) Insect and Disease Disturbances Correlate With Reduced Carbon Sequestration in Forests of the Contiguous United States. Front. For. Glob. Change 4:716582. Volume 4 Article 716582 doi: 10.3389/ffgc.2021.716582
monarch butterfly on swamp milkweed; photo by Jim Hudgins, USFWS
I have been impressed recently by two groups of scientists who are trying to broaden understanding of the impacts of invasive plants by examining the interactions of those plants with insects. As they note, herbivorous insects are key players in terrestrial food webs; they transfer energy captured by plants through photosynthesis to other trophic levels. This importance has been recognized since Elton first established the basic premises of food webs (1927) [Burghardt et al.; full citation at end of blog] Arthropods comprise significant members of nearly every trophic level and are especially important as pollinators. If introduced plants cause changes to herbivore communities, there will probably be effects on predators, parasites, and other wildlife through multitrophic interactions [Lalk et al.; Tallamy, Narango and Mitchell].
[I briefly summarize the findings of a third group of scientists at the end of this blog. The third group looks at the interaction between agriculture – that is, planting of non-native plants! – and climate change.]
One approach to studying this issue, taken by Douglas Tallamy of the University of Delaware and colleagues, is to look at the response of herbivorous insects to NIS woody plants fairly generally. They integrate their studies with growing concern about the global decline in insect populations and diversity. They note that scientists have focused on light pollution, development, industrial agriculture, and pesticides as causes of these declines. They decry the lack of attention to disruption of specialized evolutionary relationships between insect herbivores and their native host plants due to widespread domination by non-indigenous plants [Richard, Tallamy and Mitchell].
In their studies, Tallamy and colleagues consider not just invasive plants, but also non-native plants deliberately planted as crops or ornamentals, or in forestry. They point out that such introduced plants have completely transformed the composition of plant communities in both natural and human-dominated ecosystems around the globe. At least 25% of the world’s planted forests are composed of tree species not native to their locale. At least one-sixth of the globe is highly vulnerable to plant invasions, including biodiversity hotspots [Richard, Tallamy and Mitchell].
A different approach, taken by Lalk and colleagues, is more closely linked to concern about impacts of the plants themselves. They have chosen to pursue knowledge about relationships between individual species of invasive woody plants and the full range of arthropod feeding guilds – pollinators, herbivores, twig and stem borers, leaf litter and soil organisms. In so doing, they decry the general absence of data.
Both teams agree that:
Invasive plants are altering ecosystems across broad swaths of North America and the impacts are insufficiently understood.
The invasive plant problem will get worse because non-native species continue to be imported and planted. (Reminder: the Tallamy team considers impacts of deliberate planting as well as bioinvasion.)
Plant-insect interactions are the foundation of food webs, so changes to them will have repercussions throughout ecosystems.
Tallamy team
Non-native plants have replaced native plant communities to a greater or lesser extent in every North American biome – including anthropogenic landscapes [Burghardt]. The first trophic level in suburban and urban ecosystems throughout the U.S. is dominated by plant species that evolved in Southeast Asia, Europe, and South America [Tallamy and Shropshire]. Abundant non-native plants not only dominate plant biomass; they also reduce native plant taxonomic, functional and phylogenetic diversity and heterogeneity. Thus, they indirectly alter the abundance of native insects [Burghardt; Richard, Tallamy and Mitchell].
I think these articles might actually underestimate the extent of these impacts. While Richard, Tallamy and Mitchell report that more than 3,300 species of non-native plants are established in continental U.S., years ago Rod Randall said that more than 9,700 non-native plant species were naturalized in the U.S. (probably includes Hawai` i. The Tallamy team cites USDA Forest Service data showing 9% of forests in the southeast are invaded by just 33 common invasive plant species [Richard, Tallamy and Mitchell], I have cited that and other sources showing even greater extents of plant invasion in the east and here; other regions and here.
The Tallamy team has conducted several field experiments that demonstrate that the presence of non-native plants suppress numbers and diversity of native lepidopteran caterpillars. These non-native woody plants have not replaced the ecological functions of the native plants that used to support insect populations. This is true whether or not the non-native plants are deliberately planted or are invading various ecosystems on their own. [Richard, Tallamy and Mitchell]. (Of course, they expect plant invasions to grow; they note that some of the many ornamental species that are not yet invasive will become so.)
The result is disruption of the ecological services delivered by native plant communities, including the focus of their studies: plants’ most fundamental contribution to ecosystem function: generation of food for other organisms [Burghardt].
They note that plants’ relationship to insects differs depending on the insects’ feeding guilds — folivores, wood eaters, detritivores, pollinators, frugivores, and seed-eaters; and among herbivores with different mouthparts — chewing or sucking; and as host plant specialists or generalists. They decry studies that fail to recognize these differences [Tallamy, Narango, and Mitchell].
The Tallamy team explores why insect populations decline among non-native plants. That is,
1) Do insects directly requiring plant resources have lower fitness when using non-native plants; do they not recognize them as viable host plants; or do they avoid them altogether?
2) Are reductions in numbers of specialist herbivores mitigated by generalists? A decade of research shows that both specialists and generalists decline.
The team’s studies focus on lepidopteran larvae (caterpillars). Insect herbivores are both the largest taxon of primary consumers and extremely important in transferring energy captured by plants through photosynthesis to other trophic levels [Burghardt]. In addition, insects with chewing mouthparts are typically more susceptible to defensive secondary metabolites contained in leaves than are insects with sucking mouthparts that tap into poorly defended xylem or phloem fluids [Tallamy, Narango and Mitchell].
A study by Burghardt et al. found that 75% of all lepidopteran species and 93% of specialist species were found exclusively on native plant species. Non-native plants that were in the same genus as a native plant often supports a lepidopteran community that is a similar but depauperate subset of the community found on its native congener. In fact, the insect abundance and species richness supported by non-native congeners of native species was reduced by 68%.
A meta-analysis of 76 studies by other scientists found that, with few exceptions, caterpillars had higher survival and were larger when reared on native host plants. Plant communities invaded by non-native species had significantly fewer Lepidoptera and less species richness. In three of eight cases examined, non-native plants functioned as ecological traps, inducing females to lay eggs on plants that did not support successful larval development. Richard, Tallamy and Mitchell cite as an example the target of many conservation efforts, monarch butterflies (Danaus plexxipus), which fail to reproduce when they use nonnative swallowworts (Vincetoxicum species.) instead of related milkweeds (Asclepias species.).
Tallamy and Shropshire ranked 1,385 plant genera that occur in the mid-Atlantic region by their ability to support lepidopteran species richness. They found that introduced ornamentals are not the ecological equivalents of native ornamentals. This means that solar energy harnessed by introduced plants is largely unavailable to native specialist insect herbivores.
Tallamy, Narango, and Mitchell describe the following patterns:
1) Insects with chewing mouthparts are typically more susceptible to defensive secondary metabolites contained in leaves than are insects with sucking mouthparts that tap into poorly defended xylem or phloem fluids. As a result, sucking insects find novel non-indigenous plants to be acceptable hosts more often. However, there are more than 4.5 times as many chewing (mandibulate) insect herbivores than sucking (haustellate) species. It follows that the largest guild of insect herbivores is also the most vulnerable to non-native plants as well as being the most valuable to insectivores.
native azalea Rhododendron periclymenoides; photo by F.T. Campbell
2) Woody native species, on average, support more species of phytophagous insects than herbaceous species.
3) Although insects are more likely to accept non-native congeners or con-familial species as novel hosts, non-native congeners still reduced insect abundance and species richness by 68%.
4) Host plant specialists are less likely to develop on evolutionarily novel non-indigenous plants than are insects with a broader diet. There are far more specialist species than generalists, so generalists will not prevent serious declines in species richness and abundance when native plants are replaced by non-indigenous plants. In addition, non-native plants cause significant reductions in species richness and abundance even of generalists. In fact, generalists are often locally specialized on particular plant lineages and thus may function more like specialists than expected.
5) Any reduction in the abundance and diversity of insect herbivores will probably cause a concomitant reduction in the insect predators and parasitoids of those herbivores – although few studies have attempted to measure this impact beyond spiders, which are abundant generalists. The vast majority of parasitoids are highly specialized on particular host lineages.
6) Studies comparing native to non-native plants must avoid using native species that support very few phytophagous insects as their baseline, e.g., in the mid-Atlantic region tulip poplar trees (Liriodendron tulipifera) and Yellowwood (Cladrastus kentuckea).
7) Insects that feed on well-defended living tissues such as leaves, buds, and seeds are less likely to be able to include non-native plants in their diets than are insects that develop on undefended tissues like wood, fruits, and nectar. Although this hypothesis has never been formally tested, they note the ease with which introduced wood borers – emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot-hole borers, redbay ambrosia beetle, Sirex woodwasp (all described in profiles posted here — have become established in the US.
palamedes swallowtail Papilio palamedes; photo by Vincent P. Lucas; this butterfly depends on redbay, a tree decimated by laurel wilt disease vectored by the redbay ambrosia beetle
Lalk and Colleagues
As noted, Lalk and colleagues have a different frame; they focus on individual introduced plant species rather than starting from insects. They also limit their study to invasive plants. The authors say there is considerable knowledge about interactions between invasive herbaceous plants and arthropod communities, but less re: complex interactions between invasive woody plants and arthropod communities, including mutualists (e.g., pollinators), herbivores, twig- and stem-borers, leaf-litter and soil-dwelling arthropods, and other arthropod groups.
They ask why this knowledge gap persists when invasive shrubs and trees are so widespread and causing considerable ecological damage. They suggest the answer is that woody invaders rarely encroach on high-value agricultural systems and some are perceived as contributing ecosystem services, including supporting some pollinators and wildlife.
Lalk and colleagues seek to jump-start additional research by summarizing what is currently known about invasive woody plants’ interactions with insects. They found sufficient data about 11 species – although even these data are minimal. They note that all have been cultivated and sold in the U.S. for more than 100 years. All but one (mimosa) are listed as a noxious weed by at least one state; two states (Rhode Island and Georgia) don’t have a noxious weed list. None of the 11 is listed under the federal noxious weed statute.
Ailanthus altissima
Illustrations of how minimal the existing information is:
Tree-of-heaven (Ailanthus altissima) is noted to be supporting expanded populations of the Ailanthus webworm moth (Atteva aurea), which is native to Central America; and to be the principal reproductive host for SLF (Lycorma delicatua) https://www.dontmovefirewood.org/pest_pathogen/spotted-lanternfly-html/
Chinese tallow (Triadica sebifera) is thought to benefit both native generalist bee species and non-indigenous European honeybees (Apis mellifera).
Chinese privet (Ligustrum sinense) appears to suppress populations of butterflies, bees, and beetles.
Lalk and colleagues then review what is known about interactions between individual invasive plant species in various feeding guilds. They point out that existing data on these relationships are scarce and sometimes contradictory.
They believe this is because interactions vary depending on phylogenetic relationships, trophic guild, and behavior (e.g., specialized v. generalist pollinator). Arthropods can be “passengers” of a plant invasion. That is, they can be affected by that invasion, with follow-on effects to other arthropods in the community. Also, arthropods can be “drivers” of invasion, increasing the success of the invasive plants.
They then summarize the available information on various interactions. For example, they note that introduced plants can compete with native plants in attracting pollinators, causing cascading effects. Or they can increase pollination services to native plants by attracting additional pollinators.
They note that herbivore pressure on invasive plants can have important impacts on growth, spread, and placement within food webs. They note that these cases support the “enemy release hypothesis”, although they think there are probably additional driving mechanisms.
Lalk and colleagues note that most native twig- and stem-borers (Coleoptera: Buprestidae, Curculionidae, Cerambycidae; Hymenoptera: Siricidae) are not considered primary pests but that some of our most damaging insect species are wood borers (see above).
Some of these borers are decomposers; in that role, they are critical in nutrient cycling.
Arthropods in leaf litter and soil also serve important roles in the decomposition and cycling of nutrients, which affects soil biota, pH, soil nutrients, and soil moisture. They act as a trophic base in many ecosystems. Lalk and colleagues suggest these arthropod communities probably change with plant species due to differences in leaf phytochemistry. They cite one study that found litter community composition differed significantly between litter beneath tree-of-heaven, honeysuckle (Lonicera maackii), and buckthorn (Rhamnus cathartica) compared to litter underneath surrounding native trees.
Recommendations
Both the Tallamy and Lalk teams call for ending widespread planting of non-native plants. Lalk and colleagues discuss briefly the roles of
The nursery industry (including retailers); they produce what sells.
Scientists and educators have not sufficiently informed home and land owners about which species are invasive or about native alternatives.
Private citizens buy and plant what their neighbors have, what they consider aesthetically pleasing, or what is being promoted.
States have not prohibited sale of most invasive woody plants. Regulatory actions are not a straightforward matter; they require considerable time, supporting information, and compromise.
Tallamy team calls for restoration ecologists in the eastern U.S. to consider the number of Lepidopterans hosted by a plant species when deciding what to plant. For example, oaks (Quercus), willows (Salix), native cherries (Prunus)and birches (Betula) host orders of magnitude more lepidopteran species in the mid-Atlantic region than tulip poplar.(Those lepidopteran in turn support breeding birds and other insectivorous organisms.) [Tallamy & Shropshire]
Lalk and colleagues focused on identifying several key knowledge gaps:
How invasive woody plants affect biodiversity and ecosystem functioning
How they themselves function in different habitats.
Do non-native plants drive shifts in insect community composition, and if so, what is that shift, and how does it affect other trophic levels?
How do IAS woody plants affect pollinators?
The authors do not minimize the difficulty of separating such possible plant impacts from other factors, including climate change and urbanization.
Outhwaite et al. (full citation at end of this blog) note that past studies have shown that insect biodiversity changes are driven primarily by land-use change (which is another way of saying planting of non-native species – as Dr. Tallamy and colleagues describe it) and increasingly by climate change. They south to examine whether these drivers interact. They found that the combination of climate warming and intensive agriculture is associated with reductions of almost 50% in the abundance and 27% in the number of species within insect assemblages relative to levels in less-disturbed habitats with lower rates of historical climate warming. These patterns were particularly clear in the tropics (perhaps partially because of the longer history of intensive agriculture in temperate zones). They found that high availability of nearby natural habitat (that is, native plants) can mitigate these reductions — but only in low-intensity agricultural systems.
Outhwaite et al. reiterate the importance of insect species in ecosystem functioning, citing pollination, pest control, soil quality regulation & decomposition. To prevent loss of these important ecosystem services, they call for strong efforts to mitigate climate change and implementation of land-management strategies that increase the availability of natural habitats.
SOURCES
Burghardt, K. T., D. W. Tallamy, C. Philips, and K. J. Shropshire. 2010. Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities. Ecosphere 1(5):art11. doi:10.1890/ES10-00032.
Lalk, S. J. Hartshorn, and D.R. Coyle. 2021. IAS Woody Plants and Their Effects on Arthropods in the US: Challenges and Opportunities. Annals of the Entomological Society of America, 114(2), 2021, 192–205 doi: 10.1093/aesa/saaa054
Richard, M. D.W. Tallamy and A.B. Mitchell. 2019. Introduced plants reduce species interactions. Biol Invasions
Tallamy, D.W., D.L. Narango and A.B. Mitchell. 2020. Ecological Entomology (2020), DOI: 10.1111/een.12973 Do Non-native plants contribute to insect declines?
Tallamy, D.W. and K.J. Shropshire. 2009. Ranking Lepidopteran Use of Native Versus Introduced Plants Conservation Biology, Volume 23, No. 4, 941–947 2009 Society for Conservation Biology DOI: 10.1111/j.1523-1739.2009.01202.x
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm