The Nature Conservancy (TNC) and Clemson University have analyzed how to persuade people not to move firewood – and the tree pests that can accompany it. (Full citation at the end of this blog) Their study is based on five surveys conducted by TNC between 2005 and 2016. These surveys guided TNC’s “Don’t Move Firewood” campaign and its outreach efforts since the beginning in 2008
As Solano et al. note, wood-boring pests continue to enter the country and spread, causing immense damage. Firewood transport by campers is a significant contributor to that spread. Millions of individuals decide whether to move firewood. Yet the scientific literature is quite limited regarding their behavior and TNC’s survey data has never been published.
The patchwork of state and federal quarantines is largely reactive and has failed to prevent continuing spread. The regulatory regime has been further fragmented by APHIS’ deregulation of the emerald ash borer. As a consequence, limiting the spread of pests depends even more on educating campers to behave responsibly – voluntarily.
The TNC’s surveys each focused on different geographic areas and asked different questions in each. So their compilation cannot show trends in awareness or other measures. Nevertheless, the authors find:
Most people in the United States don’t know firewood can harbor invasive forest insects and diseases, but when targeted by effective education they can learn and are likely to change their behavior.
The two best ways to reach the public is through emails confirming campsite reservations and flyers handed out at parks. Web-based information seemed less effective. However, most of the surveys were done before 2011, the year when 50% of adults reported using internet media.
Forestry-related public agencies (especially state forestry departments) are the most trusted sources of information about forest health issues.
It works better to “push” information, not expect people to seek it on their own.
Messages should focus on encouraging the public to make better choices, including how they, themselves, will benefit. Positive, empowering calls to action, like “Buy it Where You Burn It” or “Buy Local, Burn Local” are better than negative messages, such as “Don’t Move Firewood”.
People respond to messages that emphasize protecting forest resources, e.g., ecosystem services like clean water. They response less to messages about forest threats.
Hungerford Lake Recreation Area at Equestrian Campground. Original public domain image from Flickr
Solano et al. describe the ways that different socioeconomic groups differ in their awareness of forest pests and in how they respond to various statements about forests, pests, and messengers. The focus is on how to overcome four psychological barriers to changing behavior that had been identified in a study of climate change. In the firewood context, those barriers were: 1) lack of awareness; 2) mistrust and negative reactions to the messengers; 3) habit; and 4) social comparison, norms, conformity, and perceived poor quality of purchased firewood.
From this work, the authors suggested further work::
Development of education and outreach programs that target those with lower education levels, since, on average, ~60% of people who camp did not graduate from college. Further research is probably needed to identify the most effective messengers and messages.
While 80% of the survey respondents were over 40, the proportion of campers made up of Gen X and millennials is increasing. Managers need to improve outreach for younger audiences. This includes engaging the messengers they trust: scientists, environmentalist politicians, peer networks, and social media.
While women trust the USDA Forest Service and conservation organizations, 55% of campers in a given year are men. Further research is needed to clarify the most effective messengers and messages for men. The outreach agencies should select the messengers that both sexes trust.
Levels of awareness should be assessed both before and after implementing new educational strategies so that the strategies’ effectiveness can be determined.
Since 80% of the respondents were white, determining the most effective messages and messengers for other ethnic groups also seems necessary, although the authors did not address this.
SOURCE:
Solano, A., Rodriguez, S.L., Greenwood, L., Rosopa, P.J., and Coyle, D.R. 2022. Achieving effective outreach for invasive species: firewood case studies from 2005-2016. Biological Invasions. https://link.springer.com/article/10.1007/s10530-022-02848-w
To request a copy of this study from the author, contact the lead author at Clemson University.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
spotted knapweed (Centaurea maculosa); photo by Alan Vernon via Wikipedia
Litt and Pearson (full citation at the end of the blog) are trying to improve scientists’ ability to predict the impact of biological invasions. Their goal is to predict which organisms will be winners, which losers, in the face of anthropogenic ecosystem change.
They focus on exotic plant invasions, because they are ubiquitous. Furthermore, plant invasions affect ecosystems by reassembling the plant community in ways that affect the niches used by native animals and hence the animals’ success under the new conditions. After determining the differences between the traits exhibited by invasive plants vs. the native plants they are displacing, scientists can then identify which native animals are most likely to be affected, as well as how and why they might respond to exotic plant invasion. [Note that Doug Tallamy is looking at similar issues.]
Litt and Pearson have developed a framework to assess how plants’ traits might affect associated wildlife. Applying the framework requires certain baseline information about the ecosystem in question.
This knowledge is applied in stepwise fashion:
1) Identify the fauna of interest and their linkage to the native plant community. This association might be food or habitat values such as shelter. Then the researcher determines the relevant plant traits of importance to that animal and approximates the strength of the animal’s dependence on these traits. Note that the focus is on plant traits relevant to the animal users, rather than specific plant species.
2) Determine overall importance of the plant traits for the area under study by (a) averaging dependence of a representative subsample of individuals to obtain a community-level value for each plant species or functional group and (b) quantifying the relative abundance of the plant functional group in the community (e.g., cover or biomass).
3) Plot the way the animal species’ abundance changes with resource abundance.
4) Understand how the invasive plants will alter the distributions of the native plants’ traits and potentially introducing novel traits that might alter the faunal community.
Litt and Pearson reviewed earlier studies to test how well this framework explained the responses of three groups of fauna to plant invasions in different ecosystems.
searching for spotted knapweed; photo by Oregon Department of Agriculture
Spiders in invaded grasslands
Intermountain grasslands of western Montana are heavily invaded; non-native plants already comprise 25–60% of average total plant cover.
One group of native spiders construct their irregular webs entirely within a single plant. A second group – orb weavers – suspend their larger webs from multiple plants. The former depend on the architectural complexity of individual plants; they can build larger webs in plant species possessing greater branching and/or longer branches of the flowering stalks. Orb spiders depend more on the complexity of the overall plant community.
Plant architecture is closely tied to the plant’s functional groups, that is, whether they are grasses or forbs.
These grasslands are generally dominated by perennial grasses. The irregular-web spiders can use grasses, but strongly favor forbs, particularly those with the most complex flowering structures. Orb weavers are generalists, incorporating multiple plant species; but they also tend to favor forbs, presumably because they are more robust.
Invasive plants in the Western Montana grasslands are of two types: an annual grass, cheatgrass (Bromus tectorum), and numerous perennial and annual forbs. Cheatgrass largely replaces the dominant native grasses with a similar architecture – although cheat is shorter. The exotic forbs, which can collectively invade at levels comparable to cheatgrass, tend to be taller and more complex structurally than the native forbs. Thus, invasion by exotic forbs strongly shifts the community-level distribution of the key trait toward greater structural complexity by replacing the dominant, but structurally simplistic, native grasses, and the more diminutive native forbs. These changes increased the abundance of both spider groups, but especially the specialist irregular web weavers. They find the new conditions meet their needs. Both spider groups appeared to expand their realized niches in response to invasion, i.e., they are able to use a broader range of plant architectures than was available in the native system.
Chaetodipus sp. photo by J.N. Stuart
Rodents in semi-desert grasslands invaded by Lehmann lovegrass
In the semi-desert grasslands of the American southwest, native grasses and forbs provide food and habitat for a variety of rodents. This vegetation influences which species of rodents are present in two ways: the size of the plants’ seeds and the density of vegetative cover. Litt and Steidl examined both. They divided the rodents into separate guilds based on diet and preferred vegetative cover. The two sets of guilds did not overlap for all species.
In southern Arizona, the native plant community is dominated by several grass species and herbaceous forbs; most species produce relatively large seeds. Vegetative cover is generally low, but varies in a patchy fashion. The rodent communities in uninvaded native grasslands are dominated by seed-eaters that prefer sparse cover.
Invasion of these grasslands by Lehmann lovegrass (Eragrostis lehmanniana) results in increased vegetative cover but the grass produces very small seeds that probably provide little to no food for rodents. Another result is a decrease in overall abundance of arthropods. The new conditions favor different rodent species from those most common in uninvaded habitat.
Two more specialized seed-eating rodent species, which seek both lower cover and larger seeds, decreased in abundance. A rodent species which favors lower vegetative cover and feeds on larger invertebrates also declined. In contrast, abundance increased for two other rodent species that prefer more dense cover and are more opportunistic in their feeding. One species surprised the scientists: Dipodomys merriami increased in abundance, despite the fact that this species favors more open environments. Perhaps other functional traits or biotic interactions are important to this species? There was no apparent change in abundance for three other species, suggesting either a lack of statistical power (2 were less abundant) or that these rodents were able to persist through a balance of positive and negative changes in food and habitat characteristics.
Lucy’s warbler [nest in saguaro, not cottonwood); photo by Dominic Sherony
Warblers in Riparian Habitats in the Southwest
Riparian habitats in the same desert region have been aggressively invaded by the exotic shrub saltcedar (Tamarix spp.). Litt and Pearson consider the findings of Mahoney et al. of this invasion’s impact on two ecologically similar warbler species. One, the yellow warbler (Setophaga petechia), is very widely distributed across North America; it is considered a generalist. The other, Lucy’s warbler (Oreothlypis luciae), is endemic to a small region of the southwest United States and northern Mexico.
The two species have similar feeding behaviors but differ in their nesting requirements. The yellow warbler constructs open cup nests in the branches of shrubs and trees. Lucy’s warbler nests in cavities in larger trees excavated by others. Hence, these species were expected to respond similarly to changes in food resources and foraging habitat, but differ in their responses to changes in nesting substrate.
Native vegetation in the region consists primarily of willows and cottonwoods in the riparian corridors, with oak and mesquite woodlands in the adjacent uplands. Saltcedar invasion rapidly displaces the willows; it takes much longer to displace cottonwoods since are large and long-lived. Upland vegetation is uninvaded and unaffected. While saltcedar is structurally similar to native willows, its leaf architecture allows more light to penetrate in saltcedar stands. This can exacerbate heat stress on nestlings in these hot, arid environments, as well as expose the nestlings to nest predation. These effects are exacerbated by the presence of a biocontrol leaf beetle (Diorhabda spp.), which cause widespread defoliation of saltcedar during nesting season. Meantime, the cavity nests used by Lucy’s warbler are barely affected.
The study by Mahoney et al. showed that in low-invasion riparian sites, the two warblers occur at comparable abundances. When saltcedar invasion replaces willows, yellow warblers decline by ~50% while there is no apparent change in abundance of Lucy’s warblers.
Litt and Pearson point out that their framework is based on two key assumptions that establish the context for its efficacy.
The first is that bottom-up forces fuel ecological processes. Plants are key to making the sun’s energy available to consumer animals and – thence to predators. Consumers’ and predators’ top-down effects are secondary. The authors’ framework thus provides better predictions of community outcomes when systems are predominantly structured by bottom-up forces. As top-down forces increase or when invasive plants differentially affect multiple dimensions of the consumer niche space, it will be more challenging to track and predict outcomes, as our rodent example demonstrates.
The second assumption is that exotic plant invasions will most strongly influence bottom-up processes. Invasive plants displace native plants and their plant traits, thus directly affecting consumers by altering the quality and quantity of food and habitat resources. However, plant community changes caused by plant invasions can also affect predators directly and indirectly via several interactions. These changes in predators’ abundance and/or their per capita effects on prey might create feedbacks that can complicate interpreting and predicting invasion outcomes.
Litt and Pearson concluded that their approach is promising but has inherent limitations linked to the dynamic nature of ecological systems.
[Ecologists continue to evaluate the impacts of saltcedar eradication efforts on another bird species, the federally endangered southwestern willow flycatcher (Empidonax extimus trailii). See, for example, Goetz, A., I. Moffit and A.A. Sher. 2022. Recovery of a native tree following removal of an invasive competitor with implications for endangered bird habitat. Biological Invasions Vol. 24, pp. 2769-2793.]
SOURCE
Litt, A.R. and D.E. Pearson. 2022. A functional ecology framework for understanding and predicting animal responses to plant invasion. Biol Invasions https://doi.org/10.1007/s10530-022-02813-7
& Supporting Information [warblers in riparian ecosystems invaded by tamarisk]
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Pinus radiata plantation in New Zealand; photo by Jon Sullivan
As countries and conservation organizations ramp up tree planting as one solution to climate change, I worry that many of the plantings will use species not native to the region – with the risk of promoting more bioinvasions. My second fear is that inadequate attention will be paid to ensuring that the propagules thrive.
Warning from New Zealand
New Zealand has adopted a major afforestation initiative (“One Billion Trees”). This program is ostensibly governed by a policy of “right tree, right place, right purpose”. However, Bellingham et al. (2022) [full citation at end of blog] say the program will probably increase the already extensive area of radiata pine plantations and thus the likelihood of exacerbated invasion. They say the species’ potential invasiveness and its effects in natural ecosystems have not been considered.
Bellingham et al. set out to raise the alarm by evaluating the current status of radiata, or Monterrey, pine (Pinus radiata) in the country. They note that the species already occupies ~1.6 M ha; the species makes up 90% of the country’s planted forests. Despite the species having been detected as spreading outside plantations in 1904, it is generally thought not to have invaded widely.
The authors contend that, to the contrary, radiata pine has already invaded several grasslands and shrublands, including three classes of ecosystems that are naturally uncommon. These are geothermal ecosystems, gumlands (infertile soils that formerly supported forests dominated by the endemic and threatened kauri tree Agathis australis), and inland cliffs. Invasions by pines – including radiata pine – are also affecting primary succession on volcanic substrates, landslides on New Zealand’s steep, erosion-prone terrain, and coastal sand dunes. Finally, pine invasions are overtopping native Myrtaceae shrubs during secondary succession. Bellingham et al. describe the situation as a pervasive and ongoing invasion resulting primarily from spread from plantations to relatively nearby areas.
kauri; photo by Natalia Volna, iTravelNZ
The New Zealanders cite data from South America and South Africa on the damaging effects of invasions by various pine species, especially with respect to fire regimes.
Furthermore, their modelling indicates that up to 76% of New Zealand’s land area is climatically capable of supporting radiata pine — most of the country except areas above 1000 m in elevation or receiving more than 2000 mm of rainfall per year. That is, all but the center and west of the South Island. This model is based on current climate; a warmer/drier climate would probably increase the area suitable to radiata pine.
These invasions by radiata pine have probably been overlooked because the focus has been on montane grasslands (which are invaded by other species of North American conifers). [See below — surveys of knowledge of invasive plants’ impacts.]
Bellingham et al. recognize the economic importance of radiata pine. They believe that early detection of spread from plantations and rapid deployment of containment programs would be the most effective management strategy. They therefore recommend
1) taxing new plantations of non-indigenous conifers to offset the costs of managing invasions, and
2) regulating these plantations more strictly to protect vulnerable ecosystems.
They also note several areas where additional research on the species’ invasiveness, dispersal, and impacts is needed.
Survey of Awareness of Invasive Plants
A few months later a separate group of New Zealand scientists published a study examining tourists’ understanding of invasive plant impacts and willingness to support eradication programs (Lovelock et al.; full citation at end of the blog). One of the invasive plant groups included in the study are conifers introduced from North America and Europe. These conifers are invading montane grasslands, so they are not the specific topic of the earlier article. The other is a beautiful flowering plant, Russell lupine. These authors say that both plant groups have profound ecological, economic, and environmental impacts. However, the conifers and lupines are also highly visible at places valued by tourists. Lovelock et al. explored whether the plants’ familiarity – and beauty – might affect how people reacted to descriptions of their ecosystem impacts.
Visitors from elsewhere in New Zealand were more aware of invasive plants’ impacts and more willing to support eradication programs for these species specifically. Asian visitors had lower awareness and willingness to support eradication of the invasives than tourists from the United Kingdom, Europe, or North America. This pattern remained after the tourists were informed about the plants’ ecological impacts. All groups were less willing to support eradication of the attractive Russell lupine than the conifers.
Conifers invading montane grasslands are perhaps the most publicized invasive plants in New Zealand [as noted above]. Lovelock et al. report that New Zealand authorities have spent an estimated $NZ166 million to eradicate non-native conifers over large tracts of land on the South Island. Still, only about half the New Zealand visitors surveyed were aware of the ecological problems caused by wild conifers.
invasive lupines in New Zealand; photo by Michael Button via Flickr
Russell lupine (Lupinus × russellii) is invading braided river systems, modifying river flows, reducing nesting site availability for several endangered birds, and provides cover for invasive predators. While initially planted in gardens, the lupines were soon being deliberately spread along the roads to ‘beautify’ the landscape. Foreign tourists often specifically seek river valley invaded by the lupine because pictures of the floral display appear in both official tourism promotional material & tourist-related social media. It is not surprising, then, that even among New Zealanders, only a third were aware of the lupines’ environmental impacts.
The oldest participants (those over 60) had the lowest acceptance of wild conifers. Participants 50–59 years old were most aware of ecological problems caused by wild conifers. Participants 30–39 years old showed the highest acceptance of wild conifers and lowest awareness of ecological issues.
Female participants showed a higher preference for the landscape with wild conifers (45.90%) than males (36.89%). Female participants were also half as aware of ecological problems (25.62% v. 46.12% among male participants).
Nearly all survey participants (96.1%) preferred the landscape with flowering lupine; only 19.4% were aware of associated ecological problems. New Zealand domestic visitors were more aware. After the impacts of lupines were explained, half decided to support eradication. However, the same proportion of all survey participants (42.5%) still wanted to see lupines in the landscape.
Once again, participants older than 50 were more aware of ecological problems arising from lupine invasions. Both men and women greatly preferred the landscape with Russell lupins.
While the authors do not explore the ramifications of the finding that younger people are less aware of invasive species impacts, I think they bode ill for future protection of the country’s unique flora and fauna. They did note that respondents had a high level of acceptance overall for these species on the New Zealand landscapes.
While the study supported use of simple environmental messaging to influence attitudes about invasive species, also showed that need to consider such social attributes as nationality and ethnicity. So Lovelock et al. call for investigation of how and why place of origin and ethnicity are important in shaping attitudes towards invasives. Conveying conservation messages will be more difficult because tourist materials often contain photographs of the lupines. Much of this information comes from informal media such as social media, which are beyond the control of invasive species managers.
SOURCES
Bellingham, P.J., E.A. Arnst, B.D. Clarkson, T.R. Etherington, L.J. Forester, W.B. Shaw, R. Sprague, S.K. Wiser, and D.A. Peltzer. 2022. The right tree in the right place? A major economic tree species poses major ecological threats. Biol Invasions Vol.: (0123456789) https://doi.org/10.1007/s10530-022-02892-6
Lovelock B., Y. Ji, A. Carr, and C-J. Blye. 2022. Should tourists care more about invasive species? International and domestic visitors’ perceptions of invasive plants and their control in New Zealand. Biological Invasions (2022) 24:3905–3918 https://doi.org/10.1007/s10530-022-02890-8
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
EAB-killed ash tree falls before it can be taken down; photo courtesy of former Ann Arbor mayor John Hieftje
Concerned by growing impacts of bioinvasion and inadequate responses by national governments worldwide and by international bodies, a group of experts have attempted to determine how much invasive species are costing. They’ve built the global database – InvaCost. See Daigne et al. 2020 here.
Several studies have been based on these data. In two earlier blogs, I summarized two of these articles, e.g., Cuthbertet al. on bioinvasion costs, generally, and Moodleyet al. on invasive species costs in protected areas, specifically. Here, I look at two additional studies. Ahmed et al. focusses on the “worst” 100 invasives affecting conservation — as determined by the International Union of Conservation and Nature (IUCN). The second, by Turbelin et al., examines pathways of introduction. Full citations of all sources appear at the end of this blog.
It is clear from all of these papers that the authors (and I!) are frustrated by the laxity with which virtually all governments respond to bioinvasions. Thus more robust actions are needed. The authors and I also agree that data on economic costs influence political decision-makers more than ecological concerns. However, InvaCost – while the best source in existence — is not yet comprehensive enough to generate the thoroughly-documented economic data about specific aspects of bioinvasion that would be most useful in supporting proposed strategies.
Scientists working with InvaCost recognize that the data are patchy. At the top level, these data demonstrate high losses and management costs imposed by bioinvasion. The global total – including both realized damage and management costs – is estimated at about $1.5 trillion since 1960. In fact, these overall costs are probably substantially underestimates (Cathbert et al.). [For a summary of data gaps, go to the end of the blog.] Furthermore, they recognize that species imposing the highest economic costs might not cause the greatest ecological harm (Moodley et al).
Comparing estimated management costs to estimated damage, the authors conclude that countries invest too little in bioinvasion management efforts and — furthermore — that expenditures are squandered on the wrong “end” of bioinvasion – after introduction and even establishment, rather than in preventive efforts or rapid response upon initial detection of an invader. While I think this is true, these findings might be skewed by the fact that fewer than a third of countries reporting invasive species costs included data on specifically preventive actions. Cuthbert et al. notes that failing to try to prevent introductions imposes an avoidable burden on resource management agencies. Ahmed et al. developed a model they hope will overcome the perverse incentives that lead decision-makers to either do nothing or delay.
Why Decision-Makers Delay
Citing the InvaCost data, the participating experts reiterate the long-standing call for prioritizing investments at the earliest possible invasion stage. Ahmed et al. found that this was the most effective practice even when costs accrue slowly. They ask, then, why decision-makers often delay initiating management. I welcome this attention because we need to find ways to rectify this situation.
They conclude, first, that invasive species threats compete for resources with other threats to agriculture and natural systems. Second, Cuthbert et al. and Ahmed et al. both note that decision-makers find it difficult to justify expenditures before impacts are obvious and/or stakeholders demand action. By that time, of course, management of invasions are extremely difficult and expensive – if possible at all. I appreciate the wording in Ahmed et al.: bioinvasion costs can be deceitfully slow to accrue, so policy makers don’t appreciate the urgency of taking action.
Cuthbert et al. also note that impacts are often imposed on other sectors, or in different regions, than those focused on by the decision-makers. Stakeholders’ perceptions of whether an introduced species is causing a “detrimental” impact also vary. Finally, when efficient proactive management succeeds – prevents any impact – it paradoxically undermines evidence of the value of this action!
Ahmed et al. point out that in many cases, biosecurity measures and other proactive approaches are even more cost effective when several species are managed simultaneously. They cite as examples airport quarantine and interception programs; Check Clean Dry campaigns encouraging boaters to avoid moving mussels and weeds; ballast water treatment systems; and transport legislation e.g., the international standard for wood packaging (ISPM#15) [I have often discussed the weaknesses in ISPM#15 implementation; go to “wood packaging” under “Categories” (below the archive list)].
pallet “graveyard”; photo by Anand Prasad
Pathways of Species’ Introduction
Tuberlin et al. focus on pathways of introduction, which they say influence the numbers of invaders, the frequency of their arrival, and the geography of their eventual distribution. This study found sufficient data to analyze arrival pathways of 478 species – just 0.03% of the ~14,000 species in the full database. They found that intentional pathways – especially what they categorized as “Escape” – were responsible for the largest number of invasive species (>40% of total). On the other hand, the two unintentional pathways called “Stowaway” and “Contaminant” introduced the species causing the highest economic costs.
Tuberlin et al. therefore emphasize the importance of managing these unintentional pathways. Also, climate change and emerging shipping technologies will increase potential invaders’ survivability during transit. Management strategies thus must be adapted to countering these additive trends. They suggest specifically:
eDNA detection techniques;
Stricter enforcement of ISPM#15 and exploring use of recyclable plastic pallets (e.g., IKEA’s OptiLedge); [see my blog re: plastic pallets, here]
Application of fouling-resistant paints to ship hulls;
Prompt adoption of international agreements addressing pathways (they cite the Ballast Water Management Treaty as entered into force only in 2017 — 13 years after adoption);
Ensuring ‘pest free status’ (per ISPM#10) before allowing export of goods—especially goods in the “Agriculture”, “Horticulture”, and “Ornamental” trades; and
Increasing training of interception staff at ports.
What InvaCost Data say re: Taxa of greatest concern to me
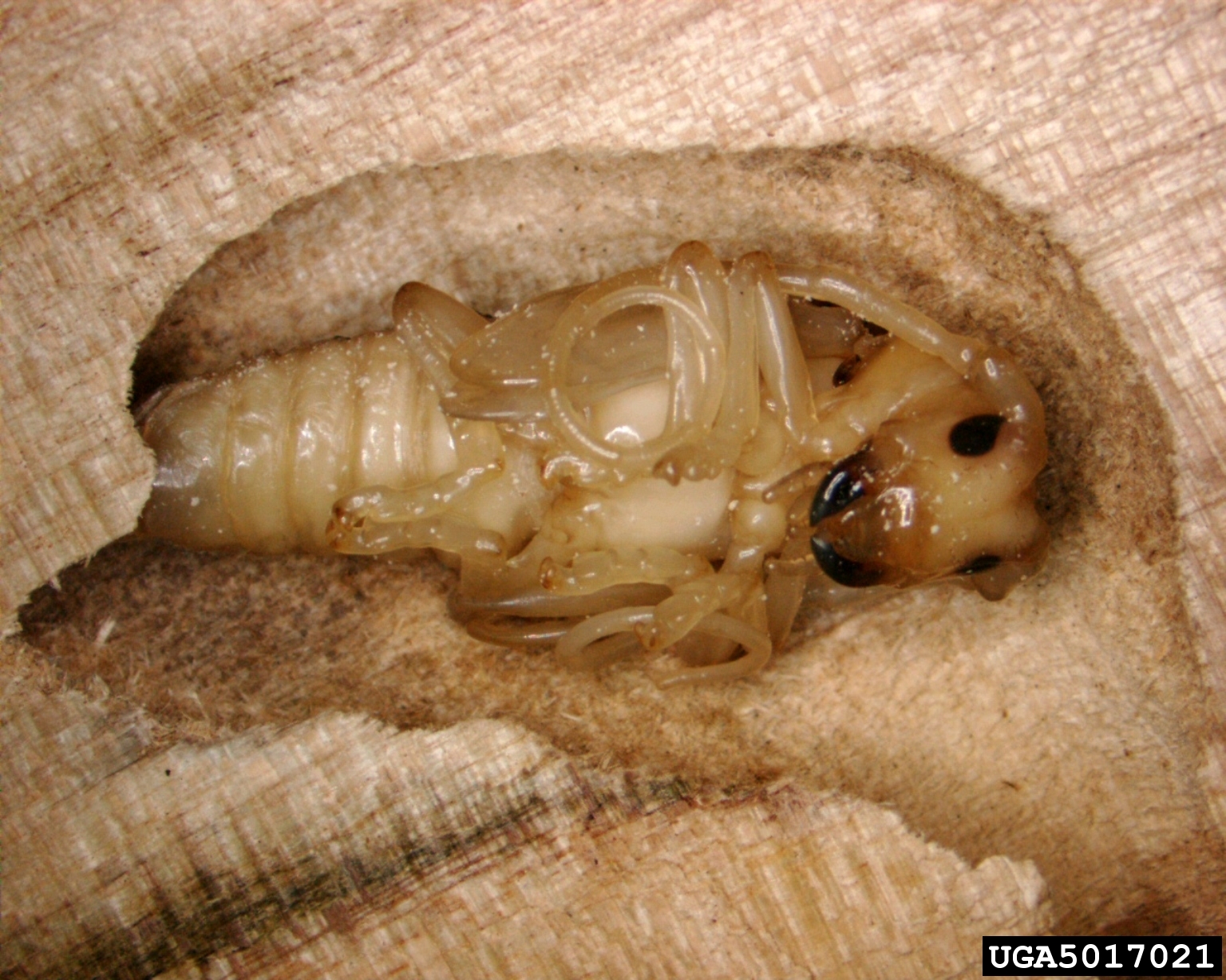
Two-thirds of reported expenditures are spent on terrestrial species (Cuthbert et al.). Insects as a Class constitute the highest number of species introduced as ‘Contaminants’ (n = 74) and ‘Stowaways’ (n = 43). They also impose the highest costs among species using these pathways. Forest insects and pathogens account for less than 1% of the records in the InvaCost database, but constitute 25% of total annual costs ($43.4 billion) (Williams et al., in prep.). Indeed, one of 10 species for which reported spending on post-invasion management is highest is the infamous Asian longhorned beetle (Tuberlin et al.)
ALB pupa in wood packaging; Pennsylvania Dept. of Natural Resources via Bugwood
Mammals and plants are often introduced deliberately – either as intentional releases or as escapes. Plant invasions are reported as numerous but impose lower costs.
Tuberlin et al. state that intentional releases and escapes should in theory be more straightforward to monitor and control, so less costly. They propose two theories: 1) Eradication campaigns are more likely to succeed for plants introduced for cultivation and subsequently escaped, than for plants introduced through unintentional pathways in semi-natural environments. 2) Species introduced unintentionally may be able to spread undetected for longer; they expect that better measures already exist to control invasions by deliberate introductions. I question both. Their theories ignore that constituencies probably like the introduced plants … and the near absence of attention to the possible need to control their spread. This is odd because elsewhere they recognize conflicts over whether to control or eradicate “charismatic” species.
Geographies of greatest concern to me
North America reported spending 54% of the total expenditure in InvaCost. Oceania spent 30%. The remaining regions each spent less than $5 billion. (Cuthbert et al.)
North America funded preventative actions most generously than other regions. Cuthbert suggests this was because David Pimentel published an early estimate of invasive species costs. I doubt it. The Lacey Act was adopted in 1905. USDA APHIS was formed in 1972 – based on predecessor agencies — because officials recognized the damage by non-native pests to agriculture. APHIS began addressing natural area pests with discovery of the Asian longhorned beetle in 1996. Of course, most of APHIS’ budget is still allocated to agricultural pests. I conclude that North America’s lead in this area has not resulted in adequate prevention programs.
Oregon ash swamp before attack by EAB (photo by Wyatt Williams, Oregon Dept. of Forestry)
Equity Issues
Tuberlin et al and Moodley et al. address equity issues of who causes introductions vs. who is impacted. This is long overdue.
More than 80% of bioinvasion management costs in protected areas fell on governmental services and/or official organizations (e.g. conservation agencies, forest services, or associations). With the partial exception of the agricultural sector, the economic sectors that contribute the most to movement of invasive species are spared from carrying the resulting costs (Moodley et al.)
A lack of willingness to invest might represent a moral problem when the invader’s impacts are incurred by regions, sectors, or generations other than those that on whom management action falls (Ahmed et al.)
People are perhaps more inclined to spend money to mitigate impacts that cause economic losses than those that damage ecosystems (Tuberlin et al.)
Data deficiencies
Only 41% of countries (83 out of 204) reported management costs; of those, only 24 reported costs specifically associated with pre-invasion (prevention) efforts (Cuthbert et al.).
Reliable economic cost estimates were available for only 60% of the “worst” invasive species (Cuthbert et al.)
Only 55 out of 266,561 protected areas reported losses or management costs (Moodley et al.).
Information on pathways of introduction was available for only three species out of 10,000 (Turbelin et al).
Taxonomic and geographic biases in reporting skew examples and possibly conclusions (Cuthbert et al.).
SOURCES
Ahmed, D.A., E.J. Hudgins, R.N. Cuthbert, .M. Kourantidou, C. Diagne, P.J. Haubrock, B. Leung, C. Liu, B. Leroy, S. Petrovskii, A. Beidas, F. Courchamp. 2022. Managing biological invasions: the cost of inaction. Biol Invasions (2022) 24:1927–1946 https://doi.org/10.1007/s10530-022-02755-0
Moodley, D., E. Angulo, R.N. Cuthbert, B. Leung, A. Turbelin, A. Novoa, M. Kourantidou, G. Heringer, P.J. Haubrock, D. Renault, M. Robuchon, J. Fantle-Lepczyk, F. Courchamp, C. Diagne. 2022. Surprisingly high economic costs of bioinvasions in protected areas. Biol Invasions. https://doi.org/10.1007/s10530-022-02732-7
Turbelin, A.J., C. Diagne, E.J. Hudgins, D. Moodley, M. Kourantidou, A. Novoa, P.J. Haubrock, C. Bernery, R.E. Gozlan, R.A. Francis, F. Courchamp. 2022. Introduction pathways of economically costly invasive alien species. Biol Invasions (2022) 24:2061–2079 https://doi.org/10.1007/s10530-022-02796-5
Williams, G.M., M.D. Ginzel, Z. Ma, D.C. Adams, F.T. Campbell, G.M. Lovett, M. Belén Pildain, K.F. Raffa, K.J.K. Gandhi, A. Santini, R.A. Sniezko, M.J. Wingfield, and P. Bonello 2022. The Global Forest Health Crisis: A Public Good Social Dilemma in Need of International Collective Action. Submitted
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
healthy hemlocks in Cook Forest, Pennsylvania; photo by F.T. Campbell
I blogged recently about North Carolina’s multi-pronged hemlock conservation program. As noted there, scientists are putting considerable hope in biological control as the most promising strategy to protect eastern (Tsuga canadensis) and Carolina hemlocks (T. caroliniana) from the hemlock woolly adelgid (HWA; Adelges tsugae). For a more detailed discussion of the adelgid’s life cycle, go here.
A new study by Crandall, Lombardo and Elkinton (full citation at end of blog) cheers us by supporting the probable efficacy of this approach – as long as a complete suite of biocontrol agents is deployed. The study points to the need to introduce additional biocontrol agents, specifically those that feed in the summer.
The study analyzed the relative importance of two different mechanisms to protect plants from herbaceous insects: do some hemlock species have an enhanced ability to fend off the adelgid (bottom-up protection); or do predators apply sufficient pressure (top-down protection) to reduce adelgid populations to levels that the tree can withstand? The study simultaneously analyzed
(1) the relative importance of summer-active and winter-active native predators;
(2) whether HWA colonization and abundances differed on western and eastern hemlock species;
(3) the relative importance of top-down and bottom-up forces on HWA feeding on western and eastern hemlocks in the adelgid’s native range;
4) tested whether the adelgid is ubiquitous at low densities across the Pacific Northwest (PNW) and compared HWA abundance in PNW to invaded range in New England.
The study was carried out in Washington State, where both western hemlock (Tsuga heterophylla) and HWA are native. They were able to compare adelgid impacts on eastern hemlock because the tree is planted in parks and gardens in the PNW.
Eastern hemlock infested by HWA; USDAFS via Bugwood
In an earlier study, (Crandall et al. 2020) found that L. nigrinus was not able to reduce HWA densities in the east. Laricobius spp have their greatest impact on HWA by larval feeding on the progrediens eggs produced by the sistens. However, 90% of hatching progrediens die naturally because there are a finite number of needles for them to settle on. To have an impact on HWA populations, Laricobius spp would have to prey on more than 90% of progrediens eggs. The solution appears to be summer-active predators – e.g., silver flies — which feed on the progrediens eggs and the sistens eggs which the progrediens generation lays.
western hemlock in British Columbia; photo by F.T. Campbell
KEY FINDINGS
Western hemlock is a native host of the adelgid. Crandall, Lombardo, and Elkinton found no evidence that western hemlock’s structure, chemistry, or other attributes help it fend off adelgid attack. The proportion of branches colonized by HWA was significantly higher on western than on eastern hemlock. Indeed, HWA populations were able to reach levels similar to those in eastern North America and were able to persist on western hemlock for multiple generations. Thus there is no evidence for bottom-up control of HWA on western hemlock.
HWA survival was significantly lower on branches of western hemlock when predators were allowed access. Crandall assumes that the smaller, non-significant, decrease in HWA densities on eastern hemlocks in the Pacific Northwest is also attributable to predation, although the data are too few to support a definitive conclusion. These predators included a species that has been released as a biocontrol agent in the east, Laricobius nigrinus. More important, apparently, was the presence of summer-active predators, including Leucotaraxis spp. and generalists. These summer-active predators are active from the progrediens nymph stage in April through the aestivating sistens nymph stage until about October. Laricobius nigrinus doesn’t become active until September. These results support the hypothesis that predator-caused mortality is responsible for suppressing HWA during rare and localized outbreaks on western hemlock in the PNW. In the east there are no native natural enemies that attack HWA – which is introduced to the region.
Effective control of HWA on the eastern naïve hosts will require establishment of a suite of predators which – together — attack the adelgid during both summer and winter.While several possible biocontrol agents have been introduced in the region, and at least some – e.g., Laricobius nigrinus – have established self-sustaining populations, are spreading, and have high predation rates, they have had very limited success in reducing HWA populations. Crandall, Lombardo and Elkinton say these data support the recent decision by the USDA Forest Service to augment the HWA biocontrol effort by introducing two species of silver flies, Leucotaraxis argenticollis and Le. piniperda, that feed on both the sistens and progrediens generations in PNW.
Tree-adelgid interactions are probably significantly affected by the lineage of both – whether the tree species has co-evolved with the specific lineage of the adelgid with which it is interacting. Crandall, Lombardo and Elkinton think evaluation of any Tsuga species’ resistance to HWA or any potential biocontrol agent needs to be studied in relation to the appropriate lineage of the adelgid.
When they compared HWA abundance (in 2021) on hemlock forests in western Washington with HWA abundance at introduced HWA range in New England, Crandall, Lombardo and Elkinton found that HWA abundance was higher in New England. They note that these comparisons are between two different linages of HWA – the lineage native to PNW and the introduced Japanese lineage in the East.
The authors note that HWA densities in the PNW are higher at the urban site (Seattle) than rural sites. Perhaps the reason is lower densities of HWA predators in non-forest settings because some, e.g., La. nigrinus, require a duff layer for pupation. Duff layers are rarely permitted to accumulate in urban areas. The authors call for studies to assess the relative abundance and identify factors affecting the abundance of HWA predators in rural and urban settings.
SOURCES
Crandall R.S., Jubb C.S., Mayfield A.E., Thompson B., McAvoy T.J., Salom S.M. and J.S. Elkinton. 2020. Rebound of Adelges tsugae spring generation following predation on overwintering generation ovisacs by the introduced predator Laricobius nigrinus in the eastern United States. Biological Control 145, 104-264. https://doi.org/10.1016/j.biocontrol.2020.104264
Crandall, R.S., J.A. Lombardo, and J.S. Elkinton. 2022. Top-down regulation of hemlock woolly adelgid (Adelges tsugae) in its native range in the Pacific Northwest of North America. Oecologia 199, 599-609. https://doi.org/10.1007/s00442-022-05214-8
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Senate Appropriations Committee has adopted its recommendations for funding APHIS and the US Forest Service in Fiscal Year 2023, which begins on October 1. The full Senate has not yet acted; most people expect that it will not act before October, so the agencies will have to operate under a “continuing resolution” for at least the first several months. Under a “CR”, funding is maintained at the current level.
SOD-infected rhododendron plants detected by state officials in Indiana in 2019
Funding for APHIS in FY23
The Senate Appropriations Committee issued a report [available here] that recognizes APHIS’ objective of protecting the animal and plant resources of the Nation from diseases and pests. These objectives are carried out through, inter alia, Safeguarding and Emergency Preparedness/Response and Safe Trade and International Technical Assistance.
The Committee recommends the following funding for specific APHIS programs (in $millions)
PROGRAM
FY22 FUNDING
FY23 ADMIN REQ
HOUSE $
SENATE COMM RECOMM
CISP ASK
Border inspections (AQI appropriated)
33.849
36.725
36.650
X
Pest Detection
28.218
29.137
29.825
29.075
30
Methods Development
21.217
21.854
31.807
23.557
23
Specialty Crops
209.533
219.533
219.698
222.072
219
Tree & Wood pests
61.217
62.854
62.562
62.719
70
Subtotal, Plant health
379.144
385.560
397.603
X
Emerg. Prepare & Response
42.021
44.242
44.317
X
Specific programs mentioned:
Northern (Asian) giant hornet eradication: $1.75 million to continue cooperation with Washington State to eradicate this pest; also to improve monitoring methods and lures, and build a rapid response platforms
sudden oak death (SOD): recognize that the EU1 and NA1 strains of this pathogen threaten Douglas-fir / tanoak forests and lead foreign governments to impose quarantines on U.S. timber exports. So APHIS should spend no less that FY22 funding to better understand threat and treatment methods in wildlands. This earmark disappoints because it focuses on APHIS’ role as certifying timber exports as pest-free rather than the spread of the pathogen within the U.S. via the nursery trade. The same language appears in the report’s discussion of the Agriculture Research Service (see below).
Pertinent action re: Agriculture Research Service
The Senate Committee report sets several priorities, including the following:
Invasive Pests: The Committee is concerned about the threats invasive pests pose to agriculture, the economy, environment, human health, and national security of the Pacific region. The Committee directs ARS to continue working with stakeholders in the region to assess options for combatting invasive species, including biocontrol research facilities, containment facilities, additional laboratory space.
Sudden oak death: the same language as for APHIS. Again, I wish the language referred to the pathogen’s spread via the nursery trade.
These numbers are disappointing; the increase for “specialty crops” demonstrates the lobbying clout of the nursery and berry industries! I appreciate the attention to sudden oak death – with the caveat I mentioned.
SOD-infected tanoaks in southern Oregon; photo by Oregon Department of Forstry
Forest Service
The Senate Appropriations Committee issued a report [available here] . The Senate Appropriations Committee recommends the following funding levels for USFS programs that address non-native forest pests and other invasive species (in $millions):
PROGRAM
FY22 FUNDING
FY ADMIN REQUEST
HOUSE $
S COMM RECOMM
CISP ASK
Research
296.616
317.733
$360.4
$302.773
317.733
State & Private Forest Health Protection TOTAL
48
59.232
$52.232
50
83
S&P FHP Federal lands
16,000
22,485
?
17,000
51
S&P FHP non-federal lands
32,000
36,747
?
33,000
32
R&D
The Senate wants to retain the current structure of five regional stations, International Institute of Tropical Forestry, and Forest Products Laboratory.
The Senate listed several research priorities. Two pertain to forest health: 1) needle pathogens, and 2) Northeastern States Research Cooperative working to sustain the health of northern forest ecosystems and biological diversity management. I am disappointed that no mention is made of the need to respond to 400 introduced tree-killing insects and pathogens.
planting to test ash trees’ resistance to emerald ash borer; photo courtesy of Jennifer Koch, USFS
S&P
The Senate Committee recommends a significant increase in S&P overall ($8 million above FY22 level), but not for Forest Health Management. This is disappointing.
The Committee is concerned about high tree mortality on National Forests due to bark beetle infestations and instructs USFS to work with states and tribes to prioritize insect prevention, suppression & mitigation projects.
The Committee expects the Forest Service and Bureau of Land Management (BLM) to continue efforts to treat sudden oak death in California and Oregon. It provides $3 million for this purpose, including for partnerships with private landowners.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm