Spruce budworm (Choristoneura fumiferana); photo by Jerald E. Dewey, USFS; via Bugwood; populations of several forest birds, including Cape May, Tennessee and Bay-Breasted warblers, become more numerous during budworm outbreaks
Now that Drs. Ziska and Aucott have educated us about the strong impact atmospheric CO2 can have on both plants and phytopagous insects, I have asked the experts whether these interactions have been incorporated in the models scientists are using to forecast pest activity in American forests as the climate changes.
The answer is no.
bay-breasted warbler; photograph by Dave Inman at Presque Isle State Park, PA; via Flickr
Dr. Bethany A. Bradley, Co-Director, Northeast Climate Adaptation Science Center at the University of Massachusetts, says empirical models of species range shifts typically only use climate and sometimes other environmental factors (like soils or topography) as predictors of potential geography. Inclusion of demographic processes like how plant growth is affected by more or less water, CO2, competition with other plants etc. would require a lot of data. It is currently impossible since there are tens of thousands of plant species interacting in the forests of eastern North America – and perhaps these factors have been analysed for only a hundred of them.
Mike Aucott points to the same difficulty: inclusion of CO2 in models of the future populations of specific plants would be difficult since the impacts vary from species to species and are compounded by other factors such as soil nitrogen levels, moisture levels, temperature, presence of competing plants, etc.
Regarding insects, Dr. Aucott thinks it is clear that some orders, such as Lepidoptera, don’t fare as well when feeding on plants grown under elevated CO2. He is not aware of efforts to model impacts of high CO2 on specific insects or even orders or feeding guilds.
juniper geometer (inchworm); Dr. Tallamy says inchworms are hairless & good tasting – so sought by birds
Dr. Ziska concurs about the difficulties. Dr. Ziska asks why there is so little funding to study these issues, especially given their probable impact on human food supplies and health – as described in his blog and an opinion piece published in Scientific American two years ago.
I hope that scientists, decision-makers, readers of this blog … maybe even the media! – take into consideration these complexities, even if they cannot be defined.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – [but do not address climate or CO2 aspects] review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Wildfire: one of the widely recognized results of climate change (The Pioneer Fire located in the Boise National Forest near Idaho City, ID began on Jul. 18, 2016 and the cause is under investigation. The Pioneer Fire has consumed 96,469 acres. U.S. Forest Service photo. Original public domain image from Flickr)
A guest blog by Michael Aucott. Mike is a retired research scientist of the NJ Department of Environmental Protection. He has also taught chemistry at the College of New Jersey. He is currently a member of the NJDEP Science Advisory Board Standing Committee on Climate and Atmospheric Sciences, and on the board of directors of the PA/NJ Chapter of the American Chestnut Foundation. If you wish to contact Mike, use the contact button on this website. You MUST include your email address; it is not recorded automatically.
Two major perturbations affect Earth and its living systems, climate change and habitat disruptions. Emerging data show that these are more closely related than previously realized; they are connected by carbon dioxide, CO2.
Climate change basics: the physics
Climate change concerns have focused on the alteration of weather and climate due to the increase in atmospheric concentrations of greenhouse gases, primarily carbon dioxide, CO2. The impact of CO2 on climate has been understood for at least 120 years. In 1896 the Nobel-Prize-winning Swedish chemist Svante Arhennius published calculations demonstrating that human emission of CO2, when combined with the positive feedback effects of water vapor, would warm the Earth (Arhennius, 1896). His equation, ΔF = α ln(C/C0), relates the change in climate “forcing” (the degree to which temperature change is forced) to the ratio of the concentration of CO2 currently in the atmosphere (C) to a previous concentration (C0). This equation is still in use today. Arhennius estimated that a doubling of CO2 would warm the Earth by about 4 degrees C. This estimate is not far off from current estimates based on much more elaborate calculations.
This warming impact is caused by the physics of CO2, water vapor, and other “greenhouse” gases. Infrared radiation causes the CO2, water, and other greenhouse gas molecules to vibrate, leading to the absorption of the energy carried by that radiation. Much of the solar energy coming from the sun is not in the infrared frequency range, so it passes through the atmosphere without being absorbed. However, when this energy is absorbed by the surfaces of the Earth and its biota, and is re-radiated as infrared radiation, it is then absorbed by greenhouse gases, warming the planet.
The amount of water vapor in the atmosphere is directly related to the atmosphere’s temperature: warm air holds more water vapor. Human activity hasn’t directly changed the concentration of water vapor in the atmosphere significantly. But by burning fossil fuels, humans have dramatically increased the atmospheric concentration of CO2 and in so doing, also indirectly increased the concentration of water vapor. Just as Arhennius predicted over 120 years ago, this increase in CO2 is warming the Earth.
Ramifications of this warming include increased heat episodes, the intensification of the hydrological cycle (greater frequency of both heavy precipitation events and of droughts), sea level rise due to the melting of land-based glaciers and the thermal expansion of ocean water, and, almost certainly, intensification of storms and an increase in extreme weather. These climate-warming- based perturbations have the potential to influence the functioning of Earth’s biota in many deleterious ways, and clearly can be associated with the many facets of habitat disruption.
Climate change amplifications: the chemistry
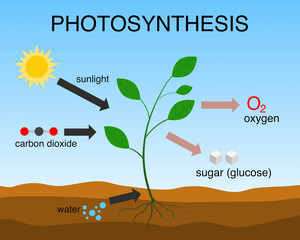
But there’s another aspect of CO2 that may be more important insofar as habitat disruption is concerned and that has been largely ignored: chemistry. CO2 is a trace gas as far as we humans and other animals are concerned, unnoticed by our bodies in normal life. But to plants it is a vital food. It is taken up by plants as an essential input to photosynthesis. In this chemical reaction, using the energy of sunlight, plants combine CO2 and water vapor to make oxygen and carbohydrates, represented with a generic formula of CH2O, according to the equation CO2 + H2O → CH2O + O2. Without this reaction, life as we know it would not exist.
The atmospheric concentration of CO2 has varied over time; some 50 million years ago it was considerably higher than today. However, for at least the last three million years, the concentration of CO2 has been in the range of 280 ppm. Over these millions of years biota have adapted to this concentration. But within the last 300 years, one ten thousandth of this period – a blink of an eye in the geological or evolutionary time scale – the concentration of CO2 has shot up to 420 ppm, a 50% increase, due to humanity’s burning of fossil fuels and forests.
Imagine what might happen to a person who had been on a tight dietary budget for most of his or her life but suddenly got access to 50% more carbohydrates, but no more protein or minerals? We could expect major changes in the metabolism of that person. This dramatic change is what has, in effect, happened to the whole of life on Earth. Our planet’s primary biota, plants, now suddenly have the opportunity to gorge on CO2. But their access to other important substances such as nitrogen has not changed. Evidence is accumulating that CO2 at its elevated level of 420 ppm is not, as has been proclaimed by some, a healthy influence but is instead throwing Earth’s ecosystem into disarray.
Much of the recent experimental evidence on the impacts of enriched atmospheric CO2 has been assembled by Lewis Ziska and presented in his new book, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It (Ziska, 2022; see full citation at the end of the blog). The findings documented in this book reveal a variety of impacts of elevated CO2. These impacts include stimulation of growth of invasive plants, decreases in the nutrient content of major crops, and changes in plants’ production of insecticidal, allergenic, and other compounds. The changing chemistry of plants may be contributing to a major die-off of insects and insect-eating animals including birds. Below are some details.
Habitat Disruptions: Stimulation of Invasive Plants
The generally accepted explanation for why some plants are invasive is that they have been introduced to new regions where their historic predators and parasites aren’t present. Without these drags on their growth, they have flourished. That some alien plants are not browsed by white-tailed deer contributes significantly to their invasiveness in Eastern North America. Other factors are clearly involved as well, including changes in the temperature regime and the availability of water and other resources such as nitrogen.
But elevated CO2 is also a factor. In recent years, techniques for measuring responses of organisms in situ under elevated CO2 conditions have been developed, making possible investigations of the impacts of CO2 concentrations that could exist in the future under otherwise relatively realistic conditions. What the actual atmospheric CO2 concentration will be in 2050 or 2100 is difficult to predict; it depends on what humanity does to control emissions. Various scenarios suggest that levels could exceed 500 ppm by 2050 and might exceed 1000 ppm by 2100 (Tollefson, 2020).
cheatgrass; photo by Jaepil Cho
One study found that the invasive weed Canada thistle, Cirsium arvense, responds more strongly to elevated CO2 than soybean, a crop that it often plagues. In a high CO2 environment, this weed’s root system grows strongly enough to make it significantly more resistant to herbicides (Ziska, et al., 2004) (Ziska, 2010). The highly invasive and dangerously flammable cheatgrass (Bromus tectorum), nicknamed “grassoline” by the U.S. Forest Service because of its propensity to intensify wildfires, also responds strongly to elevated CO2 (Ziska, et al., 2005). Also found to be boosted by enriched CO2 is yellow star-thistle (Centaurea solstitialis), considered one of California’s worst weeds. In one study (Dukes, et al., 2011) this plant grew 600% larger in elevated CO2 relative to ambient, while native plants responded much less strongly or not at all. Japanese honeysuckle, Lonicera japonica, which plagues many areas in the U.S., was found to increase in biomass by 135% at a CO2 concentration of 675 ppm while a similar native plant, coral honeysuckle (Lonicera sempervirens) increased by only 40% (Sasek & Strain, 1991). In a field study also involving Japanese honeysuckle (Belote, et al., 2004), researchers found that its above ground net production (ANPP) approximately tripled under enriched CO2 while other plants in the trial showed showed lesser increases or actual decreases.
Other plants have been found to be selectively encouraged by enriched CO2 including cherry laurel (Prunus laurocerasus), invasive in the Pacific Northwest U.S. and the U.K., (Hattenschwiler & Korner, 2002); dalmation toadflax (Linaria dalmatica), invasive in much of North America (Blumenthal, et al., 2013); honey mesquite (Prosopis glandulosa) , invasive in Australia and parts of Africa (Polley, et al., 1996); and kudzu (Pueraria lobata), which afflicts the Southeast U.S. (Sasek & Strain, 1988). Three plants invasive in China or Southeast Asia, American rope (Mikania micrantha), Creeping oxeye (Wedelia trilobata), and a morning glory species (Ipomoea cairica), were found to produce 70.3% greater biomass when grown at a CO2 concentration of 700 ppm while three corresponding indigenous plants Paederia scandens, Wedelia chinensis and Ipomoea pescaprae, produced only 30.5% more biomass (Song, et al., 2009).
yellow star thistle; photo by Eugene Zelenko
The list goes on of studies showing increased growth of some plants under enriched CO2 conditions. As more in situ investigations are undertaken, it seems likely it will become clearer that today’s enriched level of CO2 is helping some plants to become invasive.
Complexities and contradictory findings exist however. Not all plants are stimulated by enriched CO2. An important difference in the response to higher levels of CO2 is whether a plant has a C3 or a C4 photosynthetic mechanism. C4 plants contain a biochemical pump that concentrates CO2, making them more adapted to low CO2 conditions (Hager, et al., 2016). At current levels of CO2, such plants’ need for CO2 is easily met. C3 plants do not have this CO2 concentrating ability, and so higher levels boost their growth. In a broad meta-analysis of literature, the average response to elevated CO2 of 365 C3 plant species and 37 C4 plant species was noted; the response was significantly increased in C3 species but was unchanged in C4 species (Robinson, et al. 2012). One striking example of such a difference was observed in the field study noted above (Belote, et al., 2004), wherein researchers found that Japanese honeysuckle (a C3 plant) was significantly encouraged by elevated CO2 relative to other plants at the same locale. The same study found that another aggressive invader, Japanese stiltgrass (Microstegium vimineum), a C4 plant, was unaffected or even slightly inhibited relative to competing plants’ growth by elevated CO2.
One finding is widespread; most plants studied under enriched CO2 regimes show an increase in biomass and evince a higher ratio of carbon to nitrogen (C/N ratio) in their tissues and an overall decline in nitrogen concentrations than when grown under ambient conditions. Since nitrogen is a key component of protein, this change can be expected to lead to lowered protein content of critical food crops. Some impacts of this change are already well underway because of today’s elevated CO2 concentration. Changes since 1850 in the C/N ratio and in the estimated protein content of an important plant product, pollen, were discovered in a striking study by Lewis Ziska and colleagues (Ziska, et al., 2016). Using archived museum samples, these researchers determined the nitrogen content of pollen of Solidago canadensis (Canada goldenrod) going back to the 1850s. They estimated that the protein content of goldenrod pollen, a vital nutrient for North American bees, has declined in inverse proportion to the rise in atmospheric CO2, dropping from a concentration of approximately 18% in the mid-1800s to approximately 12% today. They pointed out that it is possible that bees are now unable to provide sufficient protein and other nutrients to larvae, and that one of the main reasons for bee declines is malnutrition caused by enriched atmospheric CO2. Other studies also indicate that elevated CO2 could cause lower nitrogen concentrations in plants and lead to less proteinaceous plant parts, including pollen, being available to plant-feeding insects (Hall, et al., 2005; Knepp, et al., 2007).
bumblebee on goldenrod; photo by Keila
The changing C/N ratio is almost certainly already affecting the human food supply. As documented in an extensive review of published findings (Soares, et al., 2019), elevated CO2 has a considerable impact on the accumulation of minerals and protein in plants, with many plant species showing declines in both quality and quantity of key nutrients. These changes have worrisome implications for human nutrition and may already be responsible for increasing incidences of dietary deficiency in some areas. Lewis Ziska discusses the likely impact of rising CO2 on the future human food supply in his recent post. A number of studies showing declines in protein and also other nutrients such as zinc in food crops important to humanity are also described in Ziska’s new book, Greenhouse Planet, noted above.
Habitat Disruptions: Other Changes in Plant Chemistry
Other changes in plants besides nutritional content may be driven by enriched CO2. Plants produce a variety of secondary metabolites. Most plants use the C3 mechanism; with 50% more available of a key input, some changes in these plants’ production of such chemicals can be expected. Some changes have been observed. Mohan et al. (2006) report that enriched CO2 in an intact forest system increased water use efficiency, growth, and population biomass of poison ivy (Toxicodendron radicans) and that high-CO2 plants also produced a more toxic form of the allergenic compound urushiol.
Quercus chapmanii; photo by Mary Keim at Seminole State Forest, Florida
Many of the phytochemicals plants produce function as defenses against insect predation, and changes in such production have been found to impact herbivore feeding. For example, Landosky and Karowe (2014) suggest that specialist herbivores may have to contend with more effective chemical defenses by plants under elevated CO2. Hall, et al. (2005), in a study involving several oak and one legume species in a scrub oak ecosystem in Florida (see photo above), found that 700 ppm CO2 levels led to decreased damage to plants by four of six insect groups investigated. They did not see increases in plants’ production of carbon-based secondary metabolites, including total phenolic compounds, condensed tannins, hydrolyzable tannins, cellulose, hemicellulose, and lignin however. They concluded that the primary driver of decreased insect predation under elevated CO2 was lower overall plant nitrogen levels. They stated that the decline of nitrogen levels in foliage under elevated CO2 indicated lower foliar quality and hypothesized that the reductions in insect feeding stemmed from the combined effects of nutrient limitation and increases in parasitism and predation on the nutrient-constrained insects. They further stated that although insects try to compensate for lower nutrient content of leaves by eating more, they did not see an increased portion of damaged leaves in their study. These researchers did not report measurements of terpenoid compounds however, which are reported to represent the largest class of secondary metabolites (Wikipedia, 2022). In another study (Hall, et al., 2005a) found that concentrations of condensed tannins were higher in oak leaf litter under elevated CO2, which suggests that enhanced production of insecticidal compounds or other changes to plant tissues could affect not only insects that consume living plant tissue, but also detritivores.
Robinson et al. (2012) also investigated plants’ production of secondary metabolites in their literature review. Looking at all plant groups, they found that under elevated CO2 the production of nitrogen-based secondary metabolites (e.g., alkaloids, cyanogenic glycosides, and glucosinolates) decreased by 16% while the carbon-based secondary metabolites total phenolics, condensed tannins, and flavonoids increased by 19%, 22%, and 27% respectively. Another carbon-based metabolite, terpenoids, declined by 13%. They further divided plants into grasses, shrubs, herbs/forbs, and trees and found differing responses to elevated CO2. Trees, for example, showed increased production of total glycosides and total phenolics, little change in production of total flavonoids, and a decline in the production of total terpenes. Like Hall et al., (2005), Robinson et al. found a strong and significant decrease in nitrogen concentrations under elevated CO2 for C3 plants. A decrease did not show up for C4 plants.
In addition to chemical defenses, plants have physical characteristics such as surface waxes, trichomes, secretory canals, and tissue toughness-enhancing substances such as lignin. All of these features can reduce the edibility of plants for arthropod herbivores. Robinson et al. (2012) found consistent responses to these characteristics under elevated CO2; leaf toughness and specific leaf weight increased by 11% and 18%, respectively, while specific leaf area did not show a significant change. They concluded that there is an increase in general “toughness” of leaves under elevated CO2. As did Hall et al., (2005), Robinson et al. concluded that elevated CO2 will induce changes in plant chemistry, physiology, and morphology that are likely to impact the nutritional quality of host plants for insect herbivores.
Habitat Disruptions: Changes in Plant Chemistry and Insect Decline
Numerous studies have documented a recent and dramatic decline in insect populations and discussed the probable cascading impacts of such declines through the food chain, affecting spiders, lizards, birds, and other organisms (Samways, et al., 2020; Cardoso, et al., 2020; Sánchez-Bayoa & Wyckhuys, 2019; Tallamy & Shriver, 2021). It has been argued that the main drivers of insect species declines are habitat loss and conversion to intensive agriculture and urbanization; pollution, mainly by synthetic pesticides and fertilizers; biological factors, including pathogens and introduced species; and climate change.
But a puzzling aspect is that some insect declines have been observed in nature preserves (Vogel, 2017) that presumably are not greatly affected by most of the above drivers. One example is a study spanning 27 years that found a 76% decline in flying insect biomass at several of Germany’s protected areas subject to rather low levels of human disturbance (Hallmann et al., 2017). Another study in rainforests of Puerto Rico, apparently not subject to influences such as light pollution, habitat loss, pesticides, or agriculture, reported biomass losses between 98% and 78% for ground-foraging and canopy-dwelling arthropods over a 36-year period, (Lister and Garcia, 2018). This leaves climate change as the likely culprit. But although the varied impacts of climate change, including heat episodes, drought, and other episodes of extreme weather could impact insect populations in remote as well as populated areas, the trends observed appear to far exceed the magnitude of such climate-related disturbances over the last several decades.
tent caterpillars; Shiela Brown, Public Domain Pics
Another puzzling aspect is that not all insect orders or feeding guilds seem to be equally affected. Sanchez-Bayoa & Wychuys (2019) whose article reports on a review of 73 historical reports, state that Lepidoptera, Hymenoptera and dung beetles (Coleoptera) appear to be the taxa most affected in terrestrial environments. Robinson et al. (2012) found that phloem feeders such as Homoptera respond positively to elevated CO2 while foliage feeders/Lepidoptera respond negatively. Lepidoptera were among the most impacted; relative growth rate, fecundity, and abundance all declined under high CO2 conditions, while relative consumption rate, total consumption, and development time all increased.
Most Lepidoptera are herbivorous, feeding in their larval stage, caterpillars, on plants. Caterpillars are key components of the terrestrial ecology; in most forests of the world, caterpillars consume more living leaves than all other animals combined (Janzen, 1988). Insect herbivores such as caterpillars are near the hub of most terrestrial food webs, comprising essential food for insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears (Burghardt et al., 2010). At least 310 North American bird species are known to feed extensively on caterpillars, and the majority of terrestrial birds rely on insects during reproduction and other nutrient-limited periods in the annual cycle (Narango, Tallamy & Marra, 2018). Caterpillars and moths are the largest component of nestling diets in hundreds of species of migrant bird species (Tallamy & Shriver, 2021); they are among the “little things that run the world” (Wilson, 1987).
Carolina chickadee; one of the birds Dr. Tallamy focuses on because it feeds its young on caterpillars; photo by Dan Pancamo; through Wikimedia
Habitat Disruptions and Climate Change: Connected by CO2
The apparently heightened rate of decline of insect herbivores such as caterpillars compared to some other insects, and the findings that many declines have been observed in areas relatively unimpacted by direct human influences such as light pollution, pesticides, and land-use change, point to the likelihood of a broad, perhaps ubiquitous, cause. Climate change is such a broad cause. Even broader and more ubiquitous is the main driver of climate change, CO2. Every plant in the world is constantly bathed in an enriched concentration of this gas. A conclusion seems likely: CO2 is not only causing global warming and climate change but is also affecting life on this planet in ways that, while still poorly understood, are already reducing the nutritive value of food crops, may be a significant cause of the spread of invasive plants, and may be the main driver of insect declines and the cascading impacts of such declines on insect-eating animals such as birds.
What to do?
To mitigate climate change and, as argued here, to mitigate habitat disruption, the steady rise in the atmosphere’s burden of CO2 must be halted, and then steps must be taken to lower the current concentration to a healthier level. These are not hopeless tasks. Although what has been a relentless rise in CO2 emissions at the global level continues, increases have slowed and even stopped in some parts of the world. Accelerating the development of low- and zero-carbon energy sources and encouraging energy conservation, as will be done through the U.S.’s Inflation Reduction Act, will further this progress.
More will be needed. Putting a significant and steadily increasing price on the carbon in fossil fuels is arguably the most important next step. Fossil fuels enjoy a free ride. The byproduct of their combustion, CO2, is dumped with little or no restrictions into the world’s atmosphere. A price on carbon would end this inequity. There are ways this could be done in a revenue-neutral (“fee and rebate”) manner that would avoid harm to economies and those with low- and moderate-incomes. A major step forward in pricing carbon by the European Union, a carbon border adjustment mechanism, is close to implementation. For more on this and other developments in cutting CO2 emissions, see the analyses and insights of the Carbon Tax Center and learn more about actions you can take to influence governments with Citizens’ Climate Lobby.
Not discussed here, but another stark example of habitat disruption is the increasing acidification of the world’s oceans caused by the dissolution of atmospheric CO2 in the waters. The ocean’s average pH has dropped from 8.2 to 8.1 within the last 200 years. Because pH is a logarithmic scale, this represents an increase in hydrogen ion concentration of over 25%, a change that is already threatening some marine creatures. More on this is available from many sources; e.g., Kolbert (2014).
References
Arhennius, Svante, 1896, On the Influence of Carbonic Acid in the Air upon the Temperature on the Ground, Philosophical Magazine and Journal of Science, 41, 237-276.
Belote, R., J. Weltzin, and R. Norby, 2004, Response of an Understory Plant Community to Elevated [CO2] Depends on Differential Responses of Dominant Invasive Species and Is Mediated by Soil Water Availability, New Phytologist 161, 827-835.
Blumenthal, D., V. Resco, J. Morgan, D. Williams, D. LeCain, E. Hardy, E. Pendall, and E. Bladyka, 2013, Invasive Forb Benefits from Water Savings by Native Plants and Carbon Fertilization Under Elevated CO2 and Warming, New Phytologist 200, 1156-1165.
Burghardt, Karin T., D. W. Tallamy, C. Philips, and K. Shropshire, 2010, Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities, Ecosphere 1: 1-22.
Dukes, J., N. Chiariello, S. Loarie, and C. Field, 2011, Strong Response of an Invasive Plant Species (Centaurea solstitialis L.) to Global Environmental Changes, Ecological Applications 21, 1887-1894.
Hall, M., P. Stiling, B. Hungate, B. Drake, and M. Hunter, 2005a, Effects of elevated CO2 and herbivore damage on litter quality in a scrub oak ecosystem, Journal of Chemical Ecology, 31, 2343-2356.
Hall, M., P. Stiling, D. Moon, B. Drake, and M. Hunter, 2005, Effects of elevated CO2 of foliar quality and herbivore damage in a scrub oak ecosystem. Journal of Chemical Ecology 31, 267-286.
Hallmann, C.A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., Stenmans, W., Müller, A., Sumser, H., Hörren, T., Goulson, D., de Kroon, H., 2017, More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12, e0185809.
Hattenschwiler, S. and C. Korner, 2003, Does Elevated CO2 Facilitate Naturalization of the Non-indegenous Prunus laurocerasus in Swiss Temperate Forests?, Functional Ecology 17, 778-785.
Janzen, Daniel H., 1988, Ecological Characterization of a Costa Rican Dry Forest Caterpillar Fauna, Biotropica, 20, 120-135.
Knepp, R., J. Hamilton, A. Zangeri, M. Berenbaum, and E. Delucia, 2007, Foliage of oaks grown under elevated CO2 reduces performance of Antherae Polyphemus (Lepidoptera: Saturnidae), Environmental Entomology 36, 609-617.
Kolbert, E., 2014, The Sixth Extinction, Henry Holt & Co., NY
Landosky, J., and D. Karowe, 2014, Will chemical defenses become more effective against specialist herbivores under elevated CO2? Global Change Biology, 20, 3159–3176.
Lister, B., and A. Garcia, 2018, Climate-driven declines in arthropod abundance restructure a rainforest food web, PNAS 115, E10397–E10406
Mohan, J., L. Ziska, W. Schlesinger, R. Thomas, R. Sicher, K. George, and J. Clark, 2006, Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2. PNAS 103, 9086-9089.
Narango, D., D. Tallamy, and P. Marra, 2018, Nonnative plants reduce population growth of an insectivorous bird, PNAS 115: 11549–11554.
Polley, H., H. Johnson, H. Mayeux, C. Tischler, and D. Brown, 1996, Carbon Dioxide Enrichment Improves Growth, Water Relations, and Survival of Droughted Honey Mesquite (Prosopis glandulosa) Seedlings, Tree Physiology, 16, 817-823.
Robinson, E., G. Ryan, and J. Newman, 2012, A meta-analytical review of the effects of elevated CO2 on plant-arthropod interactions highlights the importance of interacting environmental and biological variables, New Phytologist 194, 321-336.
Sánchez-Bayoa, F. and K. Wyckhuys, 2019, Worldwide decline of the entomofauna: A review of its drivers, Biological Conservation 232, 8-27.
Sasek, T. and B. Strain, 1988, Effects of Carbon Dioxide Enrichment on the Growth and Morphology of Kudzu (Puerarialobata), Weed Science 36, 28 – 36, DOI: https://doi.org/10.1017/S0043174500074415
Sasek, T. and B. Strain, 1991, Effects of CO2 Enrichment on the Growth and Morphology of a Native and Introduced Honeysuckle Vine, American Journal of Botany 78, 69-75.
Soares, J., C. Santos, S Carvalho, M Pintado, and M. Vasconceios, 2019, Preserving the nutritional quality of crop plants under a changing climate: importance and strategies. Plant and Soil 443, 1-26. https://doi.org/10.1007/s11104-019-04229-0
Song, L., J. Wu, C. Li, F. Li, S. Peng, and B. Chen, 2009, Different responses of invasive and native species to elevated CO2 concentration, Acta Oecologica 35, 128-135.
Tallamy, D. and W.G. Shriver, 2021, Are declines in insects and insectivorous birds related? Ornithological Applications 123: 1–8, DOI: 10.1093/ornithapp/duaa059
Vogel, G., 2017, Where Have All the Insects Gone? Surveys in German nature preserves point to a dramatic decline in insect biomass. Key members of ecosystems may be slipping away, Science 356, 376-379.
Wilson, Edward O., 1987, The Little Things That Run the World (The Importance and Conservation of Invertebrates), Conservation Biology, 1, 344-346.
Ziska, L., 2010, Elevated Carbon Dioxide Alters Chemical Management of Canada Thistle in No-till Soybean, Field Crops Research 119, 299-303.
Ziska, L., Shaun Falukner, and John Lydon, 2004, Changes in biomass and root: Shoot Ratio of Field-grown Canada Thistle (Cirsium arvense), a Noxious, Invasive Wed, with Elevated CO2: Implications for Control with Glyphosate, Weed Science 52, 584-588.
Ziska, L.H., J.S. Pettis, J. Edwards, J.E. Hancock, M.B. Tomecek, A. Clark, J.S. Dukes, I. Loladze, and H.W. Polley, 2016, Rising atmospheric CO2 is reducing the protein concentration of a floral pollen source essential for North American bees, Proc. R. Soc. B, 283, 20160414, http://dx.doi.org/10.1098/rspb.2016.0414
Ziska, Lewis, 2022, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It, Columbia University Press, NY.
photo courtesy of Oregon Department of Agriculture
As we know, wood packaging (SWPM; crates, pallets, spools, etc.) is a high-risk pathway for introduction of bark- and wood-infesting insects (borers). (To see my 40 earlier blogs about wood packaging material, scroll down below archives to “Categories,” click on “wood packaging”.) Examples of highly damaging pests introduced to North America include Asian longhorned beetle; emerald ash borer; redbay ambrosia beetle; sirex woodwasp; possibly the polyphagous and Kuroshio shot hole borers; Mediterranean oak borer; and dozens of others. (As of 2014, 58 new species of non-native wood- or bark-boring insects had been detected in the past 30 years [Leung et al. 2014]).
The Asian longhorned beetle and emerald ash borer were probably introduced before the World Trade Organization (WTO) came into effect in 1994; many of the others were detected – if not introduced – after that date. This global trade agreement not only facilitated rapid growth in trade volumes; it also imposed stringent conditions for adoption of plant health (= phytosanitary) measures aimed at preventing pest introductions. (For a review of the WTO restrictions, see my Fading Forests II report, here).
While the risk of pests travelling in raw wood was well known, U.S. and international phytosanitary agencies became aware that wood packaging fit into that category with detection of the ALB in New York and other wood-borer introductions. They acted remarkably rapidly to reduce this risk by negotiating and adopting International Standard for Phytosanitary Measures (ISPM) #15 in 2002.
The goal of ISPM#15 is to “significantly reduce” [not eliminate] the risk of pests associated with solid wood used for constructing packaging (e.g., crates, pallets), from being introduced to other countries through international trade.
This first international standard addressing a pathway of introductions was adopted 20 years ago. (The U.S. fully implemented ISPM#15 in 2006; see either article by Haack for a description of the phase-in period.) So – how great is the risk of pest introduction in wood packaging now? What proportion of these incoming containers are likely to be harboring tree-killing insects? Since it is impossible to reduce that risk to 0 while continuing trade using wood packaging, what is an acceptable level of risk? In determining that level, we must keep in mind the huge volumes of wood packaging being used in international trade, and the serious damage these wood-borers can cause. (See the pest profiles in the links provided above.)
I applaud the international phytosanitary community for acting fast and for choosing a pathway standard rather than try to differentiate the level of risk associated with any particular transaction – given that wood packaging could be made from dozens or hundreds of tree taxa, there are thousands of species of wood-boring insects, and the likelihood of an introduction depends in part on the exporting and importing countries. Plus, international trade involves huge volumes of goods. According to Haack et al. (2022), ~ 55 million TEU (shipping containers measured as twenty-foot equivalents) entered the U.S. in 2020. This is a 68% increase over the volume in 2003. Imports in the first half of 2020 were down because of the COVID epidemic. They then grew rapidly through the first half of 2022; imports from Asia in the first 10 months of 2022 were 21% higher than in the same period in 2019 (Mongelluzzo 2022). Haack et al. (2022) note that the number of countries from which SWPM originated more than doubled from 2003–2004 to 2010–2020, although it dropped after 2018.
In 2014, Haack et al. published an estimate of the pest approach rate in wood packaging as of 2009. Depending on which countries were included and how the time periods were selected to separate pre- and post-adoption of ISPM#15, they reported a 36–52% reduction in the SWPM infestation rate following ISPM#15 implementation. This resulted in an estimated infestation rate of 0.1% (1/10th of 1%). In a recent blog, I applied this estimated approach rate to find that probably 11,000 containers per year transported pests to North America in 2021; 80% of these shipments came to the United States.
Since 2009, traders have gained 13 more years of experience. More important, in 2009 the standard was changed to require that wood packaging be constructed from wood that had been debarked before treatment. There is a tolerance limit for small patches of residual bark. Given that bark provides shelter both for insects already there, and facilitates any new infestation after the treatment was performed, it was expected that this change would further reduce the pest risk.
Since more than a decade has passed since the original analysis, and wood-borers continue to be found in wood packaging – in the U.S. and elsewhere – Haack and colleagues have re-analyzed the pest approach rates (see Haack et al. 2022). Their objectives were to
(2) compare the borer detection rates individually for three kinds of imports and key US trading partners;
(3) see whether wood borer presence varies by season; and
(4) assess the diversity of borer taxa detected overall, and by cargo category and country of origin.
Over the entire 17-year period 2003 – 2020, 87,571 consignments met the conditions for the study: they contained wood packaging that bore the ISPM#15 mark (from 2006 and onwards) indicating it had been treated as required; and the shipment was not from Canada (the U.S. does not require wood packaging from Canada to comply with ISPM#15).
They analyzed the data for the entire 17-year period and separately for four phases:
1) before the U.S. implemented ISPM#15 (2003-2004);
2) phasing-in of U.S. implementation (2005 – 2006);
3) full implementation – but without any restriction on THE presence of bark (2007-2009); and
4) full implementation with restrictions on bark (2010 – 2021).
Over the period 2003– 2020, wood borers were detected in 180 of the 87,571 consignments, or 0.21%. This was 38% less than the 0.34% infestation rate in 2003-2004, before the U.S. implemented ISPM#15. Still, the US had required China to treat its wood packaging as of December 1998 because of introduction of ALB. However, the reduction was greatest in the first phase (2005-2006); in subsequent periods the pest approach rate inched back up. Detection rates have been relatively constant since 2005 despite the requirement in 2009 that bark be removed and a resulting reduction in the presence of bark (it fell from 40% or more of inspected consignments before 2009 to 15% after 2010).
Unfortunately, the data used in the study do not indicate if borers detected on wood were located under any bark that was present. There might be some indication from the species detected: 100% of Scolytinae identified to genus or species detected before 2007 were true bark beetles (which develop primarily under bark), but only 23% in 2010–2020 period.
The data revealed no strong seasonal pattern.
Types of Wood Packaging
The study findings indicate that crates are the type of wood packaging most likely to be infested by insects. While crates constituted only 7.5% of the wood packaging inspected, they made up 29.4% of the infested packaging – or four times greater than their proportion of the dataset. Pallets constituted 88.6% of the inspected wood packaging, but only 67.2% of the infested shipments. Dunnage and “other” wood packaging made up insignificant proportions of both total wood packaging inspected and wood packaging found to be infested. (Of course, dunnage can still pose a threat; see my blog about issues in Houston with dunnage bracing breakbulk cargo.) The Haack et al. (2022) study did not examine dunnage accompanying breakbulk shipments.
Records of Various countries
The 180 infested consignments originated from 30 countries. For two of these countries, the percentage of wood packaging found to be infested was higher than the proportion of all wood packaging from that country that was inspected. Packaging from China made up 4.6% of all shipments inspected, but 22% of the 180 consignments with infested wood packaging. Thus the proportion Chinese consignments with infested wood is five times greater than expected based on their proportion of the dataset. The rate of wood packaging from China that is infested has remained relatively steady – as I noted above. The Chinese infestation rate was 1.26% during 2003–2004, and ranged from 0.58 to 1.11% during the next three periods.
I remind you, again, that the U.S. has required treatment of wood packaging from China since December 1998. Why does this country continue to ship pest-infested wood packaging to the United States? Why are the responsible agencies in the United States not taking action to correct this problem? (DHS Bureau of Customs and Border Protection enhanced its enforcement in 2017; see my blogs.)
A second country with a record of non-compliant wood packaging – Italy – has done better. The level of pest detection still exceeded their expected proportional level – that is, Italy constituted 12.7% of all inspected shipments, but had 15% of infested consignments. Still, Italy has reduced detection rates by almost two-thirds over the 17 years of the study. The Italian statistics would have been even better if there had not been a spike of infested wood in 2015 – 2018 – for unknown reasons.
The data indicate that a third country, Mexico, has improved the pest-free quality of wood packaging accompanying it exports.
Wood from Costa Rica and Turkey has deteriorated as regards pest infestation rates.The borer detection rate on Costa Rican shipments rose from 0.072% during all of 2003–2009 to 0.665% during 2010–2020. Pest-detection rates for Turkey were actually 0 during 2003–2004 (only 59 consignments) but rose to 1.05% during 2010– 2020.
Disturbing Trends
The data reveal other trends that I find disturbing:
While the pest approach rate has fallen since U.S. implementation of ISPM#15, the extent of the decline has progressively decreased during each period studied: the reduction during 2005–2006 was 61%; during 2007–2009, 47%; during 2010-2020 only 36%.
The 2010 – 2020 pest approach rate was calculated at 0.22%. This is more than double the rate based on 2009 data (0.1%, as stated in the 2014 paper). However, we should be cautious in making this comparison because the 2014 and 2022 studies used different methods (see below). The bottom line, however, is that the approach rate remains too high, in my view. Our forests continue to be exposed to the risk of introduction of highly damaging wood-boring pests. Furthermore, since the number of countries sending us infested wood packaging has increased, those potential pests include insects from a greater variety of countries (biomes).
Given the higher number of countries involved and rising proportion of wood that is infested, it is not surprising that the diversity of wood borers found in wood packaging increased. Cerambycidae were consistently the most commonly intercepted borers – making up just under half of the total for the 17 years. Scolytinae were consistently second, at 39%. Still, all major families of borers had been intercepted throughout the period.
Explanation
From the perspective of protecting our forests, what matters is whether the “current” infestation rate is significantly below the rate before ISPM#15 was implemented. As noted, the infestation rate in the 2010-2020 period (0.22%) is, on first glance, more than twice as high as the 2009 approach rate as calculated in the 2014 paper (0.1%). However, the earlier calculation excluded reports of wood packaging from China and Mexico for reasons given in the 2014 paper. Since these countries are among the top three sources of imports to the U.S., and all have had relatively high levels of infested wood packaging, this difference must have had a significant impact on the final finding.
Indeed, the supplementary materials in Haack et al. (2022) show just such a big impact. When records from China and Mexico are excluded from the calculation, the 2010-2020 approach rate appears to have been even higher — 0.272%. This is a reduction from the pre-ISPM#15 approach rate (0.299%) of only 9% — a quarter of the reduction found when data from China and Mexico were included (note the 36% reduction noted above). This difference in approach rate estimates reflects Mexico’s success in cleaning up its wood packaging (as noted above). Since China had “steady” infestation rates throughout, adding or dropping China had less of an impact.
The data do not show a significant drop in pest approach rates during the period 2010-2020 compared to pre-ISPM#15 levels, which is disappointing. Scientists do not know why this happened. It could reflect many of the reasons discussed in the 2022 paper. Perhaps the most important factor is that reporting data on a consignment basis does not allow us to detect whether the numbers of a pest species present have decreased. [See point 5 below.]
The fact is that a pallet or crate bearing the ISPM#15 mark has not proved to be reliable as to whether the wood is pest-free. (This might be because the wood had not been treated, or that it was, but the treatment failed). All the pests detected in study (after 2006) were in wood packaging bearing the ISPM#15 mark. I have noted in past blogs [click on the “wood packaging” category to bring up blogs about wood packaging and enforcement] that Customs and Border Protection also reported that nearly all the wood packaging in which that they detected insect pests bore the mark.
Conclusions: Haack et al. (2022)
Haack et al. (2022) note that U.S. imports have risen 68% by volume from 2003 to 2020 (with additional growth since; see above); however, borer detection rates have remained rather steady. This, plus the apparently lower number of woodborers established in recent years, suggest that ISPM#15 is helping to mitigate risks. However, the reduction in detection rates is less than hoped. They discuss ten possible explanations. Six of these factors were discussed in the original analysis (Haack et al. 2014); four others are new.
(1) Pest Thermotolerance. Can pests tolerate the heat treatment schedule mandated by ISPM#15? Haack et al. (2022) note that this schedule was based on one intended to kill the pinewood nematode and that it was recognized that some pests might be able to tolerate those conditions (Haack et al. 2014). The authors review the literature and conclude that some of the live borers found in heat-treated wood packaging in the study probably did survive the heat treatment. They note that studies are now under way to test temperatures that are lethal to various borers. I have raised the issue that standards must be based on lethal temperatures that can be achieved in practice; otherwise, they won’t protect forests from introduced pests.
(2) Unintentional non-compliance. The authors concluded that accidental treatment failures are likely. They note that the International Plant Protection Convention (IPPC) has issued guidance on handling and testing during heat treatment and fumigation.
(3) Fraud. The authors conclude that fraud is possible, but that the incidence at the global scale is unknown. Each country is responsible for their own compliance. Unfortunately, there is no effective means for independently testing whether treatments have been applied. Still, we note that all live insects evaluated in this paper were in wood package that bore the required stamp and was apparently compliant.
(4) Post-treatment colonization. Haack et al. (2022) note that adoption of the bark requirements in 2009 was intended to reduce re-infestation risk. They note that fewer true bark beetles (that develop under bark) have been detected in recent years compared w/ ambrosia beetles (that develop in wood). As I noted above, the survey data do not reveal whether insects detected by inspectors were under any remaining bark.
(5) AQIM data collection protocols. The authors note that reporting of wood borer detections by consignment conceals the per-piece infestation rate. There might be many fewer individuals of a pest in a container now – and this is important because fewer individuals pose a lower establishment risk (lower propagule pressure).
(6) Pre-ISPM actions. Some countries had begun requiring treatment of wood packaging before 2003, when data collection for the study began. Thus the approach rate might have already been reduced before ISPM#15 was implemented in the U.S., leading to a smaller apparent change.
(7) Level of detection. All the analyses assumed that the detection abilities of port inspectors remained the same over the 17 years of the study. However, inspectors might have improved their efficacy through improvements in training, inspection techniques, or technology. If so, the apparent impact of ISPM#15 would be lessened in recent years. Haack et al. (2022) say estimating the effectiveness of inspections is not possible in the absence of a “leakage survey” conducted on inspected goods to see how often target pests are missed.
(8) Changing trade partners. Countries have varying levels of effort and efficacy in enforcing ISPM#15.
(9) Varying trees and their associated borers. Countries and global regions are home to different tree species and associated insects. Therefore, changes in trading partners – or forest pest conditions within a country – can affect the number and species of potential pests harbored in the wood packaging approaching our borders.
(10) Practical limits on compliance. Reducing infestation levels to near zero through reliance on application of the ISPM#15 standard would require nearly universal compliance by industry, using highly effective treatments. Haack et al. (2022) note that such compliance levels might be difficult to achieve without either very strong incentives or intensive oversight and significant penalties for noncompliant exporters. I note that I have urged the U.S. to enhance both; link to blogs at least CBP has taken action to step up enforcement.
Haack et al. (2022) call for improved education and outreach by the IPPC, plus greater cooperation and information sharing among trading countries. I note that the Cary Institute is pursuing opening data on treatment facilities’ records so importers can hire the best.
Haack et al. (2022) conclude that ISPM#15 has resulted in marked decreases in rates of borer detection in wood packaging. However, problem areas remain re: some types of commercial goods and exporting countries. Given the enormous and growing volume of international trade, the relatively low risk associated with individual crates or pallets still poses a real risk for pest intro.
Still, they consider that the near global acceptance of ISPM#15 indicates a strong commitment by the world community to minimize movement of wood pests in SWPM through international trade.
Haack et al. (2022) call for several improvements. Some concern data to support analysis of the risk level. First, recording the numbers of infested pieces instead of reporting only consignments would help determine the numbers of insects reaching our borders. They also wish to learn whether when bark is present if it exceeds the current tolerance limits; and the type of treatment applied to each infested piece of wood packaging.
They also note opportunities to improve ISPM#15 implementation and enforcement through training on applying treatments, marking and repairing wood packaging, compiling the required records, and inspecting treatment facilities.
Oregon ash swamp; photo by Wyatt Williams, Oregon Department of Forestr
Faith’s Conclusions
In my view, it is less important whether the current approach rate is exactly 0.22% or somewhat less or more. What is important:
the pest approach rate is not acceptable given the huge and rising volume of imports, potential for introductions from new trading partners (with different insect faunas), and the great damage caused by wood-boring insects.
long-standing enforcement problems have not been resolved (i.e., Chinese wood packaging). Perhaps DHS CBP’s enhanced enforcement will bring improvements. CBP staff expressed disappointment in August 2022.
American government agencies must take more effective action to ensure that trade partners comply with ISPM#15. They should also look more aggressively at other actions to curtail introductions via this pathway, e.g.,
U.S. and Canada refuse to accept wood packaging from foreign suppliers that have a record of repeated violations – whatever the apparent cause of the non-compliance. Institute severe penalties to deter foreign suppliers from taking devious steps to escape being associated with their violation record.
APHIS and CBP and their Canadian counterparts provide guidance to importers on which foreign treatment facilities have a record of poor compliance or suspected fraud – so they can avoid purchasing SWPM from them. I am hopeful that the voluntary industry program described here will help importers avoid using wood packaging from unreliable suppliers in the exporting country.
Encourage a rapid switch to materials that won’t transport wood-borers. Plastic is one such material. While no one wants to encourage production of more plastic, the Earth is drowning under discarded plastic. Some firms are recycling plastic waste into pallets.
The two articles by Haack et al. – 2014 and 2022 – fully describe the methodology used, the structure of USDA’s Agriculture Quarantine Inspection Monitoring (AQIM) program, detailed requirements of ISPM#15, the phases of U.S. implementation, etc. Also see the supplemental data sheet in Haack et al. (2022) that compares the methods used in each analysis.
SOURCES
Haack RA, Britton KO, Brockerhoff EG, Cavey JF, Garrett LJ, et al. (2014) Effectiveness of the International Phytosanitary Standard ISPM No. 15 on Reducing Wood Borer Infestation Rates in Wood Packaging Material Entering the United States. PLoS ONE 9(5): e96611. doi:10.1371/journal.pone.0096611
Haack RA, Hardin JA, Caton BP and Petrice TR (2022) Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Frontiers in Forests and Global Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ. 2014. doi:10.1890/130311
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Guest blog by Lewis Ziska, Associate Professor, Environmental Health Sciences at the Columbia University
[Dr. Ziska has spent his career analyzing the impacts of CO2 and climate change on plants – and therefore on people. He served as Project Leader for global climate change at the International Rice Research Institute; then spent 24 years at the USDA’s Agricultural Research Service, where he worked primarily on documenting the impact of climate change and rising carbon dioxide levels on: Crop selection improves production; Climate and agronomic pests, including chemical management; Climate, plant biology and public health impacts on food security with a focus on nutrition and pesticide use.]
No question you’ve heard the term, “Climate Change” or “Global Warming”, or my personal favorite, “Global Weirding”. The consequences are talked and discussed in the media—as they should be—but often the media, like many Americans, is focus challenged. Or in more polite terms, they have the attention span of a hummingbird on crack. Which is to say, that simple physical consequences, like sea level rise (heat melts ice!), and stranded animals on ice (Poor polar bear!), or intense storms (newscaster whipped about in the rain, yelling to be understood) are repeated, over and over again. Understandable, makes for good TV.
But it also makes you feel separate from what is happening, these consequences of climate change are to the “other”. I don’t live near the ocean, I don’t interact with polar bears; sure we have storms, but I live in the Midwest, in one of those states that begins with a vowel. Shoot, I commute to work, try and make ends meet, I’m not some damn tree hugger. Why should I care?
To understand why, you need a bit more background, some science that isn’t always available on TV or social media when it comes to global weirding.
First, while you may not be a tree hugger, you do, in fact, interact with nature. Several times. Every day. We call those times, “breakfast”, “lunch” and “dinner”.
dinner; photo by davylin via Flickr
You depend on nature for food. And clothing. And paper. And medicine. And oxygen. And construction materials (wood), and many, many other things. So, if nature gets hinky, and the climate becomes uncertain, it might be worth your while to think about climate change, or global weirding, in a different light. What I want to do here then, is to illuminate two examples that I hope will help you see why climate could affect you, directly and significantly.
Let’s begin with plants. Those green living things that comprise the bulk of the natural world (literally, if you were to weigh the natural world, 97% would be plants, 3% animals). Then let’s look at them through two different lenses—how will climate weirding alter your food; shoot, how will it alter the air that I breathe?
Let’s start with a basic food, rice. Obviously you don’t want to mess with its production, or its nutritional quality. But that is exactly what global weirding is doing.
Rice has flowers. Not big showy ones, but flowers none the less—ones that get fertilized with pollen, and seed is produced. The seed that feeds some two billion people– or about a quarter of the earth’s population.
Like all living things, plants are heat sensitive, and for rice, and many crop plants, the degree of sensitivity varies, depending on the part of the plant in question. Take a look at the table. The crops that are listed, including rice, are the core of what the world eats. Now notice the difference in temperature sensitivity. Vegetative parts of the plant, leaves and stems, are reasonably tolerant of higher temperatures, but flowers are not. Pollen, the plant equivalent of animal sperm, is highly temperature sensitive, and if the temperatures get into the high 90s (37-38oC), they become deformed, and the rice plant doesn’t produce seeds. Same for a number of plants, ones necessary to feed 8 billion people.
Crop
Opt. Temp. Vegetative
Opt. Temp. Flowering
Failure Temp.Flowering
Maize
28-35oC
18-22oC
35oC
Soybean
25-37oC
22-24oC
39oC
Wheat
20-30oC
15oC
34oC
Rice
28-35oC
23-26oC
36oC
Sorghum
26-34oC
25oC
35oC
Cotton
34oC
25-26oC
35oC
Peanut
31-35oC
20-26oC
39oC
Data are adapted from Hatfield et al., 2011.
Doubtful you’ve seen this climate threat to the global food supply on TV or a streaming service. I caught a glimpse once of temperature and agriculture on a CNN newscast, but with the “expert” calmly stating that we would just have to grow our corn in Canada, ha-ha. (Somehow, at least for rice, it’s hard to imagine India, one of the world’s largest rice producers, moving its rice production northward to the Himalaya’s, but I digress.)
Food is fundamental. If production, especially that of a global staple like rice, is impacted by rising temperatures there will be consequences. Rising prices, reduced availability, and wide-spread hunger.
But there is more to consider. Given the global dependence on rice, any change in its nutritional quality will also have effects, especially on poorer countries that rely heavily on rice as a major food source. And here we need to delve a little deeper into another aspect of climate weirding that doesn’t make it to the popular media—that rising carbon dioxide (CO2), the primary greenhouse gas, can also directly influence plant nutrition. The reasons are complicated, but in simple terms all living things consist of elements, carbon, nitrogen, phosphorous, sulfur, copper, etc., etc. A plant gets it’s carbon from the air (CO2), but everything else (nitrogen, potassium) from the soil.
And there is an imbalance. In the last 50 years, atmospheric CO2 has increased by about 30%, and is projected to increase another 50% by the end of the century. With more CO2, plants are becoming carbon rich, but nutrient poor. Nutrient poor, because while CO2 has increased in the air, nutrients in the soil have not kept pace. A perverse carb loading at the plant level.
As a consequence, rice, and many other plants, are shifting their chemistry. For example, there is a general decline in protein, in part because protein requires nitrogen. There are similar ubiquitous declines in iron and zinc, important micro-nutrients needed for human development.
Such nutritional degradation is of obvious global importance, and does, on occasion, show up on basic media when warming / weirding is mentioned, but you’d be hard pressed to find it.
Let’s move our light to another hidden bit of science. How plants can influence the air we breathe.
As humans, we like to trade things. And a large percentage of what we trade are living organisms, from fish to trees. But what began as local, regionalized trading has grown with the global population and the needs of that population—a population of 1.6 billion at the beginning of the 20th century is now ~8 billion at the beginning of the 21st. And we haven’t stopped trading. Biological trade is not inherently bad, but it represents a historically unprecedented global movement of DNA across continents, across countries, regions, towns, cities and ecosystems. And some of the DNA, when introduced, can do great harm to the environment, the economy and to human health. That harm has a name, “Invasive Species”.
Let us focus on one such plant species introduced to Eastern Europe, one that almost every American has personal experience (ACHOO!) come fall. The species is common ragweed. An invasive plant whose introduction and spread in Eastern Europe—introduced accidently through imported seeds or contaminated hay – has resulted in enormous environmental and economic losses in agriculture and public health in recent decades. In Hungary, the most important ambient biological air pollutant is: ragweed.
collecting ragweed pollen under different climates (Author’s photo)
The photo is from studies that I led looking at how ragweed pollen would respond to temperature and carbon dioxide. (If you’re curious, ragweed likes both.) Warmer temperatures, earlier Springs, later Autumns can extend its pollen season; not only extend, but increase the amount of pollen being generated. There is even some data suggesting that rising CO2 can alter pollen chemistry, making it more allergenic (REFS). Sadly, ragweed pollen doesn’t appear as temperature sensitive as that of rice, or other agricultural plants.
I wish I could say that ragweed was the exception among allergenic plants, but it’s the rule. Parthenium weed is a highly invasive species that has spread to more than 40 countries around the world. Like ragweed its pollen are highly allergenic, but it can also produce severe rashes, like poison ivy, and is known to be poisonous to livestock. It is highly aggressive, and arriving in a new location (where it has no natural enemies) can dominate landscapes, reducing biodiversity. And as with ragweed, high temperatures, longer growing seasons, heatwaves and droughts are expanding its range, and for that matter, make controlling its spread more difficult.
Such responses among invasive species will have direct impacts on air quality, especially among those (myself included) who suffer from seasonal allergies. Gasping for air is never fun.
Estimates are that pollen and seasonal asthma affects more than 24 million of us, including 6 million kids. And yet, when watching news reports of climate change, how many times have you seen a report on pollen and air quality? On increasing allergies or asthma? Once? Twice?
I could go on, (and if you need more examples, read “Greenhouse Planet”, my latest book). But my point is this: Not all of the consequences of rising carbon dioxide and climate change, warming, weirding, whatever, make for “good” TV. There is so much more to explore. So, do yourself a favor. Take a deeper dive, find out what is happening behind the scenes.
Because if we are going to rise to the challenge, we need to know what we are fighting against. Right now, the media is exemplary on showing some things, but silent on much else of importance. Watching news coverage of climate change is a bystander watching a cataclysm, and thinking, “Boy, glad I’m not experiencing THAT!”. Yet in the simplest and most basic of terms, you are, or will be, affected– from food choices to nutrition, even your allergies. And so much more.
It isn’t just about polar bears. It’s about you. Read, Understand, Act.
A special issue of the journal Forests (Vol. 13 2022) seeks to improve understanding of the root causes of exacerbated threats from insect pests. The issue contains 15 papers; most focus on geographic areas other than North America. The journal is open access!
Choi and Park (full citations below) link increased pest risk to climate change and increased international trade. They provide brief summaries of all 15 papers. My focus here is on two articles that provide updates on the status of the emerald ash borer (EAB Agrilus planipennis) in Russia and Ukraine. The article by Davydenko et al. also examines interactions between EAB and the invasive pathogen Hymenoscyphus fraxineus, which causes ash dieback disease. In other blogs I will look at insects linked to North America (both species from North America that threaten forests on other continents, and species in Russia that pose a threat to North America) and at the overall Russian experience.
I blogged about EAB invasion of Russia in April 2021 so this is an update.
Musolin et al. (2022) (full citations below) remind us that the EAB invasions of North America and Russia were detected almost simultaneously: in Michigan and Ontario in 2002 and in European Russia (Moscow) in 2003. They conclude that both invasions probably originated from a common source (most probably China). They date the introduction to the late 1980s or early 1990s; pathways might have been wooden crafts, wood packaging, or ash seedlings. Nate Siegert used dendrological studies to estimate a similar introduction date for the North American invasion.
European ash (Fraxinus excelsior) specimen in Belgium; photo by Jean-Pol Granmont
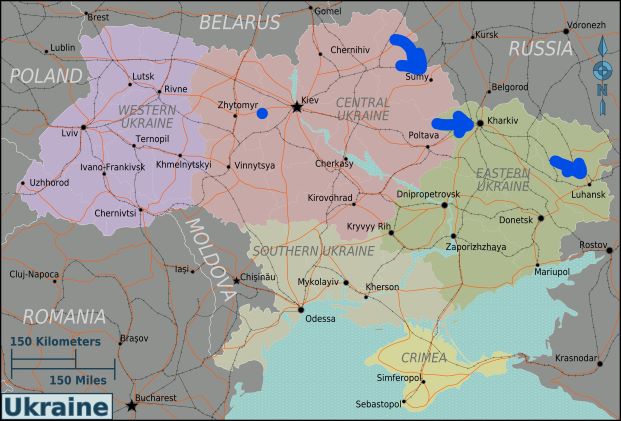
EAB has spread far in the intervening 30 + years. By early 2022, outbreaks were recorded in five Canadian provinces, 35 US states, 18 provinces and several cities in European Russia, and two provinces in Ukraine (Musolin et al. 2022) Davydenko et al. report that EAB had also established in eastern Belarus, but provide no details.
As demonstrated in the earlier blog and confirmed by Musolin et al. (2022) and Davydenko et al., the EAB has spread much faster to the southwest than directly West and to the Northwest. Davydenko et al. attribute the slower spread in the St. Petersburg area to the colder and wetter climate of this region – which is ~1200 km north of Ukraine. While the EAB reproduces in two cohorts in Eastern Ukraine, to the north the beetle requires more than one year to complete its life cycle, at least two years in the St. Petersburg area. In 2021, Musolin et al. 2021 speculated that pressure by the native parasitoid Spathius polonicus Niezabitowski might also be slowing EAB’s spread in the North. In 2022, Musolin does not address this possibility. (I note that APHIS has approved two Spathius species as biocontrol agents in the U.S.).
Musolin et al. (2022) and Davydenko et al. agree that the EAB poses real threat to ash in central and western Europe. In both the south (Davydenko et al.) and in the northwestern area around St. Petersburg ash grows in continuous stretches, linking Russia or Ukraine to Romania, Hungary, Slovakia, and Poland. These ash consist of both natural woodlands, and extensive plantings of both one of the European ash species, F. excelsior and the highly-susceptible North America green ash (F. pennsyvanica). Furthermore, the EAB is an excellent hitchhiker on vehicles & railway cars. Davydenko et al. also consider the beetle to be a strong flyer. Musolin et al. (2022) cite a separate analysis in stating that EAB can probably invade most European countries. Only some regions of Norway, Sweden, Finland, Ireland, and Great Britain are probably protected by their low temperatures.
Both articles were written too early to consider how the current war in the relevant area of Ukraine might affect spread of the EAB, although we know Ukrainians are cutting firewood. The war has certainly interrupted monitoring and other efforts.
The sources agree on EAB’s severe impacts. Musolin et al. (2022) notes that the beetle has killed millions of trees in the forests and urban plantings in North America, European Russia, and Eastern Ukraine. Davydenko et al. note that the Fraxinus genus is one of the most widely distributed tree genera in North America. They then assert that the EAB could virtually eliminate it. I know that North American scientists agree that the beetle threatens many species in the genus; but do they agree that the genus would be “virtually eliminated”? Davydenko et al. think the EAB could pose similar threat to Euro ash F. excelsior.
Musolin et al. 2022 estimate that potential economic losses in Europe could reach US$1.81 billion. By this indicator, the species ranks fourth among the most “costly” invasive pests. Russia spent an estimated US$258.9 million between 2011 and 2016.
areas of Ukraine where studies conducted
Species’ varying vulnerability
Musolin et al. (2022) cite experience in the Moscow Botanical Garden as showing that only two Asian species — Chinese ash, F. chinensis, and Manchurian ash, F. mandshurica — are were resistant to the EAB. The beetle killed both North American ash (i.e., F. pennsylvanica and F. americana) and European ash (i.e., F. excelsior, F. angustifolia, and F. ornus).
Experience in the field in Ukraine (Davydenko et al.) suggests that F. excelsior is less vulnerable to EAB than F. pennsyvanica. The overwhelming majority of EAB infestations were found on the American species. Furthermore, although similar densities of EAB larvae were found in colonized branches of both species, the proportion of larvae that were viable was significantly higher on F. pennsyvnica (91.4%) than on F. excelsior (76.1%). However, the reverse was found in the Moscow and St. Petersburg regions. Davydenko et al. don’t address directly whether they think this discrepancy is attributable to climatic factors or to differences in vulnerability between trees growing in native forests vs. human plantings. They did note that all observed cases of infestation of the native F. excelsior in Ukraine occurred in artificial plantings rather than in natural woodlands.
Davydenko et al. studied parts of Eastern Ukraine where EAB was entering areas already infected by the invasive ascomycete fungus Hymenoscyphus fraxineus (cause of ash dieback, ADB). [Two of these regions — Luhansk and Kharkiv – have been the very center of the current war.] Other studies have shown that ~1 to 5% of F. excelsior trees exhibit some resistance to ADB. These trees are thus a potential foundation for future propagation and restoration of ash in Europe – if enough of them survive attack by EAB.
They found that F. excelsior is more resistant to EAB than F. pennsylvanica, but more susceptible to ADB.
The Luhansk and Kharkiv regions have both EAB and ADB; the Sumy region has only the pathogen. EAB probably invaded the Luhansk region by 2016 (although it was detected only in 2019). The proportion of ash trees (both native and introduced species) infested rose from ~ 10–30% in 2019 to 60 – 90% by 2020–2021. The EAB arrived later in the Kharkiv region, to the Northwest, but the proportion of infested trees was similar by 2021. Combining the two regions, 75% of F. pennsylvanica trees were EAB-infested, whereas only 31% of F. excelsior trees were.
Frequencies of infections by ADB were the reverse. Pooled data from all three study regions showed 21% of F. pennsylvanica trees were infected vs. 42% of F. excelsior. In the plots invaded by both EAB and ADB (in Luhansk and Kherson regions), 4%of F. pennsylvanica were affected by both invasive species vs. 14% of F. excelsior trees. Davydenko et al. conclude that ADB facilitates EAB attack on F. excelsior trees
The impact of EAB is seen in the fact that overall mortality rates were higher in F. pennsylvanica despite the fact that in the Sumy region mortality rates were higher in F. excelsior because of the disease (EAB was absent from this region). On the other hand, EAB infests and kills F. pennsylvanica trees regardless of their prior health condition (i.e., regardless of presence/absence of ADB).
Still, fewer than half the F. excelsior trees in sites affected by both EAB & ADB (in Luhansk and Kherson regions) have died. Davydenko et al. think the survivors constitute a source of material for eventual propagation. These trees need to be carefully mapped – a task certainly not facilitated by the war!
Davydenko et al. conclude that
1. Invasion of EAB in Ukraine occurred 2–3 years before detection in 2019 [I think this is actually quite prompt for detection of EAB invasions]; the invasion is currently expanding both in terms of newly infested trees and invaded geographic area.
2. Fraxinus excelsior (at least when growing in the interior of forest stands) is more resistant to EAB than F. pennsylvanica (when growing in field shelterbelts).
3. Fraxinus excelsior is more susceptible to ADB than F. pennsylvanica.
4. Infection by ADB is likely to predispose F. excelsior to infestation by EAB.
5. Ash trees infected by ADB are predisposed for the colonization by ash bark beetles Hylesinus spp. [I did not discuss these data.]
6. Inventory and mapping of surviving F. excelsior, affected by both ADB and EAB, is necessary to acquire genetic resources for the work on strategic, long-term restoration of devastated areas, thereby tackling a possible invasion of EAB to the EU.
I was surprised that Musolin et al. (2022) think EAB’s host shift from local Asian ash species to introduced North America ash planted in the Russian Far East and China triggered EAB outbreaks in Eastern China that contributed to the beetle’s introduction to North America and European Russia. American scientists apparently agree — Haack et al. (2022) refer to both this episode and a similar to one posited for Asian longhorned beetle (Anoplophora glabripennis) — that widespread planting of Populus plantations led to rapid expansion of ALB in northern China, and the pest-weakened wood was then used in wood packaging.
Davydenko, K.; Skrylnyk, Y.; Borysenko, O.; Menkis, A.; Vysotska, N.; Meshkova, V.; Olson, Å.; Elfstrand, M.; Vasaitis, R. Invasion of emerald ash borer Agrilus planipennis and ash dieback pathogen Hymenoscyphus fraxineus in Ukraine-A concerted action. Forests 2022, 13, 789.
Haack RA, Hardin JA, Caton BP and Petrice TR (2022) Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Musolin, D.L.; Selikhovkin, A.V.; Peregudova, E.Y.; Popovichev, B.G.; Mandelshtam, M.Y.; Baranchikov, Y.N.; Vasaitis, R. North-Westward Expansion of the Invasive Range of EAB, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) towards the EU: From Moscow to Saint Petersburg. Forests 2021, 12, 502. https://doi.org/10.3390/f12040502
Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive insect pests of forests and urban trees in Russia: Origin, pathways, damage, and management. Forests 2022, 13, 521.
Siegert, N.W. 2006. 17th USDA Interagency Research Forum on Gypsy Moth and Other Invasive Species. Annapolis, MD. January 10-13, 2006.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The U.S. Geological Survey (USGS) has published an updated register of introduced species in the United States. The master list contains 14,700 records, of which 12,571 are unique scientific names. The database is divided into three sub-lists: Alaska, with 545 records; Hawai`i, with 5,628 records; and conterminous (lower 48) United States, with 8,527 records.
The project tracks all introduced (non-native) species that become established, because they might eventually become invasive. The list includes all taxa that are non-native everywhere in the locality (Alaska, Hawai`i, or 48 conterminous states) and established (reproducing) anywhere in that locality.
Each record has information on taxonomy, a vernacular name, establishment means (e.g., unintentionally, or assisted colonization), degree of establishment (established, invasive, or widespread invasive), hybrid status, pathway of introduction (if known), habitat (if known), whether a biocontrol species, dates of introduction (if known; currently 47% of the records), associated taxa (where applicable), native and introduced distributions (when known), and citations for the authoritative source(s) from which this information is drawn.
The 2022 version is more complete re: plant pathogens than earlier iterations; I thank the hard-working compilers for their efforts!
Hawai`i
wiliwili tree (Erythrina sandwicensis); photo by Forest and Kim Starr
Among the non-native species listed as being in Hawai`i are 3,603 Arthropods, including the following about which I have blogged:
eight species of mosquito in the Hawaiian islands, including the Culex and Aedes species that vector the diseases that have caused extinction of numerous endemic bird species on the Islands.
Also listed are 95 mollusk species and 20 earthworm species. I wonder who is studying the worms’ impacts? I doubt any is native to the Islands.
The Hawaiian list contains 1,557 non-native plant species. Families with largest representation are Poaceae (grass) – 223 species; Fabaceae (beans) – 156 species; and Asteraceae – 116 species. About a third of the plant species – 529 species – are designated as “widespread invaders”. This number is fifteen times higher than the numbers in lists maintained by either the Hawaiian Ecosystems At Risk project (106 species) [HEAR unfortunately had to shut down a decade ago due to lack of funds]; or Hawaiian Invasive Species Council (80 species). Furthermore, some of the species listed by HEAR and HISC are not yet widespread; the lists are intended to facilitate rapid responses to new detections. We always knew Hawai`i was being overrun by invasive species!
Among the 529 most “widespread invaders” are the following from the most introduced families:
Other families have fewer introduced species overall, but notable numbers of the most widespread invaders:
Euphorbiaceae – 8 spp. of Euphorbia
Cyperaceae – 6 spp. of Cyperus
Myrtaceae – Melaleuca quinquenervia, 2 Psidium, Rhodomyrtus tomentosa rose myrtle, 3 Syzygium [rose myrtle has been hard-hit by the introduced myrtle rust fungus]
Zingiberaceae – 3spp. Hedychium (ginger)
Anacardiaceae — Schinus molle (Peruvian peppertree); USGS considers congeneric S. terebinthifolia to be somewhat less widespread.
Plus many plant taxa familiar to those of us on the continent: English ivy, privet, castor bean, butterfly bush, Ipomoea vines … and in more limited regions, Japanese climbing fern Lygodium japonicum.
Rhus sandwicensis; photo by Forest and Kim Starr
I learned something alarming from the species profiles posted on the HISC website: the Hawaiʻi Division of Forestry and Wildlife and Hawaiʻi Department of Agriculture are considering introduction of a species of thrips, Pseudophilothrips ichini, as a biocontrol agent targetting S. terebinthifolia. I learned in early 2019, when preparing comments on Florida’s proposed release of this thrips, that Pseudophilothrips ichini can reproduce in low numbers on several non-target plant species, including two native Hawaiian plants that play important roles in revegetating disturbed areas. These are Hawaiian sumac Rhus sandwicensis and Dodonea viscosa. The latter in particular is being propagated and outplanted in large numbers to restore upland and dryland native ecosystems. While the environmental assessment prepared by the USDA Animal and Plant Service says the thrips causes minimal damage to D. viscosa, I am concerned because of the plant species’ ecological importance. Of course, the two Schinus species are very damaging invasive species in Hawai`i … but I think introducing this thrips is too risky. [To obtain a copy of CISP’s comments, put a request in comments section. Be sure to include your email address in your comment; the section algorithm does not include email addresses (how inconvenient!).]
Continental (lower 48) states
Among the 8,500 species listed in the USGS Register for the 48 continental states are 4,369 animals, among them 3,800 arthropods; 3,999 plants; and just 89 fungi. Among the arthropods, there are 1,045 beetles and 308 lepidopterans. The beetles listed include 12 Agrilus (the genus which includes emerald ash borer and goldspotted oak borer.) It does not include the elm zig-zag sawfly USGS staff have not found any publications documenting its U.S. occurrences. Among the microbes are six Phytophthora (P. cinnamomi, P. lateralis, P. pseudocryptogea, P. quercina, P. ramorum, P. tentaculata). Profiles of several of these species are posted at www.dontmovefirewood.org; click on “invasive species”, then scroll using either Latin or common name.
elm zig-zag sawfly; photo by Gyorgy Czoka via Bugwood
Citation:
Simpson, Annie, Pam Fuller, Kevin Faccenda, Neal Evenhuis, Janis Matsunaga, and Matt Bowser, 2022, United States Register of Introduced and Invasive Species (US-RIIS) (ver. 2.0, November 2022): U.S. Geological Survey data release, https://doi.org/10.5066/P9KFFTOD
United States Register of Introduced and Invasive Species;US-RIIS ver. 2.0, 2022
If you would like to contribute to future versions of the US-RIIS, please email the project leaders at us-riis@usgs.gov.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
carnation (Dianthus caryophyllus) photo by Noordzee23 via Wikimedia; carnations have been big sellers for 250 years
Plants sold in nurseries directly influence urban landscapes by providing gardens and other habitats that support humans and birds, insects, and other organisms. Doug Tallamy, though, has described ways that non-native plants fall short in providing habitat for native wildlife. Of course, non-native plants also indirectly influence natural landscapes by acting as a major source of invasive species. [see blog – includes links to regional invasive plant lists; and here] Imported plants also can carry non-native insects and pathogens – about which I blog frequently! To review these blogs, scroll down below the archives to the “categories” section and click on “plants as pest vectors”.
Now Kinlock, Adams, and van Kleunen (full citation at the end of this blog) have published a new paper that sheds more light on these issues. They analyzed the ornamental plants sold in US nurseries over 225 years (from 1719 to 1946). Their database, drawn from an earlier publication by Adams (see Sources at end of blog), included records of 5,098 ornamental vascular plant species offered for sale by 319 US nursery catalogs published over this period.
They note that present-day urban yards in the continental United States are planted in a diverse array of plants and the plants are predominately non-native species. Also, there is relatively little variation in species planted from one region to another, especially when compared to regional variation in natural areas). These patterns reflect the history of US horticulture.
Seventy percent (3,587) of the 5,098 ornamental vascular plant species offered by the 319 nurseries over those 200 years were non-native to the continental United States. They believe that the number of non-native species offered for sale has probably continued to increase in the 70 years since their study ended. They cite a study showing that 91% of tree species sold by nurseries in southern California during the 20th and early 21st centuries were not native to that state. A similar figure comes from a study of cultivated plants grown in Minneapolis–Saint Paul. There 66% of plants were non-native. (Kinlock, Adams, and van Kleunen note that 70% of species cultivated in yards of five British cities are non-native. In contrast, only 23% of cultivated plants in 18 Chinese cities were non-native.)
Kinlock, Adams, and van Kleunen note that two examples of non-native plants that have become invasive were among most common species available from nurseries beginning in the mid-19th Century: Japanese honeysuckle (Lonicera japonica) was available in 78 nurseries, and Japanese barberry (Berberis thunbergia) in 46 nurseries.
Japanese honeysuckle; photo by Chuck Bargeron, Bugwood
Historical Trends
The earliest commercial horticulture in colonies that became the United States was in the mid-17th Century. It involved imports of Eurasian fruit trees to establish orchards to provide familiar foods. Ornamental horticulture became popular earlier than I expected. Prince Nurseries was established in 1732 in Flushing, NY. It was followed by additional nurseries in New York, Philadelphia, and Massachusetts. Originally these businesses imported Old World nursery stock and seeds – again to provide familiar foods and take advantage of relationships with European contacts.
Nurseries proliferated in the 1820s in the population centers of the Atlantic coast. As people of European ancestry moved west, so did nurseries. Kinlock, Adams, and van Kleunen point out an interesting aspect of these changes: proliferation of both was aided by technology: steamboats, canals, highways, and improved mail service. Before 1800, nearly all nurseries were in the Mid-Atlantic, New England, and South. Nurseries appeared in the Great Lakes region by the 1830s. Expansion of rail lines connected nurseries from coast to coast by the 1870s. By 1890, there were more than 4,500 nurseries across the continent.
California, Florida, and Oregon are now the states with the most horticultural operations and sales (as of 2019).
The types of plants offered for sale proliferated throughout the 19th Century. The species richness of US nursery flora peaked in the early 20th Century. It decreased in the 1925 – 1946 period, possibly attributable to some combination of war-related interruptions to trade and a shift in gardeners’ focus away from ornamentals to vegetables. Another factor was adoption of international and interstate phytosanitary regulations in the early 20th Century. The post-World War II economic boom led to a new diversification of US nursery flora. In one study, a Los Angeles nursery experienced the largest increase in species richness during 1990–2011. They believe this increase was probably matched across the country. Global plant collection and importation mediated by US botanical gardens and nurseries remain active.
planting of Eucalyptus seedlings in California during 1980s; National Archives photo
Over time, nursery floras in the various regions became more similar to each other. The floras of Mid-Atlantic and New England nurseries differed before 1775, then became similar. Nurseries in the Great Lakes region also shifted toward offerings in neighboring regions. Later, nurseries in the South and West also began offering a higher proportion of species commonly sold across the continent. The nursery floras of Great Lakes and Great Plains regions were consistently similar. Still, the flora in Western nurseries still retain some unique aspects. California is the only state with a Mediterranean climate. Nurseries there sought adapted plant species, especially from an entirely new source — Australasia. (The authors note that Acacia and Eucalyptus genera, while important in California horticulture, are invaders in Mediterranean zones worldwide.) One might expect the need for plants in the Southwest to be drought-tolerant would prompt a unique nursery flora. However, the ubiquity of irrigation since the late 19th Century has blunted this necessity. Still, nursery flora in the desert biome had the most phylogenetic uniformity. The article does not discuss pressure to choose xeriscapes or otherwise adjust to current water shortages.
Pinus mogu – sought for xerescapes; photo by Krysztof Ziamk Kenraiz via Wikimedia
Growing Importance of Non-Native Species – Especially from Asia
Kinlock, Adams, and van Kleunen define “native” species as those native to the state in which it is sold; “adventive” species as native to the continental United States but not the specific state; and non-native or alien species as not native to the continental United States.
Applying these definitions, the proportion of native species in nursery flora has been consistently around 30-40% — except during the American Revolution. It rose to 70% in catalogs or advertisements published from 1775 to 1799. The authors do not speculate whether this reflected jingoism or interruptions in trade. The proportion of plant species that were adventive was 4% in the earliest period, then rose to 13% with improved transportation.
A large proportion of the native species offered in the late 18th and early 19th centuries were grown for export to Europe (think John Bartram).
Rhodendron maximum; sent to Europe by John Bartram (& invasive in Great Britain and Ireland!); photo from Pl@ntNet.identify
Throughout the 19th and 20th centuries, plants from new regions of the world with unique genetic lineages became increasingly available. Until the mid-19th Century, most non-native plants came from Europe and Eurasia. Beginning in 1850, plants native to temperate Asia composed an increasing percentage of non-native nursery flora. In the period 1900 – 1924, 19% of the ornamental nursery flora originated from temperate Asia. By the next period, 1925 – 1946, this percentage rose to 20.8%.At the same time, North American species (including some from Mexico, Canada, or Alaska) composed 21.9% of the nursery flora. (see graph).
% of species from various origins; North America – medium blue; temperate Asia – dark pink; Europe – tan; Eurasia – fuscia; Southern America – blue-green; Africa – yellow-green; Americas – olive
Plants from East Asia were particularly desirable for both biological and social reasons. First, because of climatic similarities between the two regions, East Asian plants thrived in the eastern United States. Second, popular ornamental genera had higher species richness in East Asia. Important social or cultural factors were a growing fascination with Japanese and Chinese-style gardens: forced “opening” of access to those countries in the 1840s and 1850s; and plant collecting expeditions sponsored by British and American institutions and private collectors. In 1898, the US Department of Agriculture established the Section of Seed & Plant Introduction; its purpose was to collect and cultivate economically useful non-native plants from throughout South America and Asia.
As I noted above, diversity of species in nursery offerings reached a peak in the first years of the 20th Century, concurrent with the first wave of US-sponsored plant collections; indeed, 70 species that were first listed after 1911 in their dataset were introduced by the USDA introduction program between 1912 -1942.
Commodore Perry in Japan; Library of Congress
Counter-pressures and Counter-measures
There were counter pressures during this period that – as mentioned above—probably contributed to a decline in plant introductions in later years. In the 1890s, several US states began requiring inspection of imported plant materials (spurred by plant disease outbreaks caused by spread of San Jose scale from California).
Congress adopted the Plant Quarantine Act in 1912; USDA implemented it through stringent regulations issued in 1919 (Quarantine-37). I have already noted interruption of trade associated with WWI and WWII. Kinlock et al. don’t mention the Great Depression that intervened, but I think it played a role, too. On the other hand, Q-37 was relaxed to target particular species or regions based on pest risk analysis. The article says the relaxation began in the 1930s, but I believe it actually was during the 1970s; see Liebhold et al. 2012. I have blogged several times about how well the current regulations – including the “NAPPRA” program – prevent introductions of invasive plants or damaging plant pests. To review these blogs, scroll down below the archives to the “categories” section and click on “plants as pest vectors”.
dogwood anthracnose; photo by Robert Anderson, USFS; via Bugwood
SOURCES
Adams, D.W. 2004. Restoring American Gardens: An encyclopedia of heirloom ornamental plants. Timber Press
Kinlock, N.L., D.W. Adams, M. van Kleunen. 2022. An ecological and evolutionary perspective of the historical US nursery flora. Plants People Planet. 2022;1–14. wileyonlinelibrary.com/journal/ppp3
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm