red deer on farm in New Zealand; photo by Bernard Spragg via Flickr
Among the non-native species damaging forest systems are mammals – introduced deer, goats and sheep, and swine, … These animals have the greatest impacts on island systems that are sufficiently isolated that they have no native terrestrial mammals, e.g., Hawai`i and New Zealand. Several New Zealanders have published a study of their impacts (Allen et al.; full citation at end of the blog). The focus of their analysis is the native forests’ ability to sequester carbon and thus mitigate climate change. The scientists are well aware, however, that forests provide many other ecosystem values and services, including biodiversity, water supply and quality, etc.
Introduced ungulates can have many direct effects: reduction and damage to understory biomass, depletion of seedling regeneration, exacerbated soil erosion, and local nutrient imbalances. Mammals’ browsing can modify the composition of plant communities by favoring abundance of unpalatable species. Changes also can alter ecosystem functions associated with nutrient cycling, e.g., by reducing nutrient returns to the soil and altering rates of litter decomposition
In these ways, introduced ungulates exert long-term impacts on forests’ capacity to store carbon.
Allen et al. aimed to determine the extent of these effects on forests’ capacity to store carbon, both above- and below-ground, and on forest structure and diversity. The authors compared data from 26 pairs of sites across New Zealand – half with ungulate exclosures and half adjacent unfenced control plots. The ungulate exclosures had all been established for at least 20 years. All the sites were in species-rich communities of conifers and broadleaved evergreen angiosperm trees. These forests (1) cover about one-third of the country’s remaining mature natural forest; (2) contain tree species of a wide range of palatability to ungulate herbivores; and (3) have been named a conservation priority for forest carbon management. The ungulates present on the plots were European red deer (Cervus elaphus), fallow deer (Dama dama), sika deer (Cervus nippon), and feral goats (Capra hircus).
They assert that New Zealand is a good place to do this type of study because ungulate introductions are relatively recent so their impacts are well documented.
Allen et al. found that managing invasive ungulates makes valuable contributions to conserving biodiversity but not to carbon sequestration. They found little difference in total ecosystem carbon between ungulate exclosures and unfenced control plots. Most of the difference they did find was explained by the biomass of the largest tree within each plot. As they point out, these large trees have been unaffected by invasive ungulates introduced during the last 20–50 years. However, they believe ungulate-caused changes in understory biomass, species composition, and functional diversity might result in major shifts in the diversity and composition of regenerating species. Hence, longer term consequences for both ecosystem processes and storage of forest carbon storage can be expected.
Indeed, excluding ungulates did increase the abundance and diversity of saplings and small trees. The basal area of the smallest class of tree size was 70% greater. Species richness of small trees and saplings was 44% and 68% higher, respectively. This difference had little impact on overall carbon storage, however, because the small trees and saplings store only about 5%. In contrast, the largest tree size class (dbh =/>30 cm), with their roots, contributed 44% of total ecosystem carbon in both exclosure and control plots. The largest effects of exclosures on carbon stocks were in early successional stands, e.g., those affected by such major disturbances as windthrow, volcanic activity, or landslides.
Climate change is expected to cause surprising interactions among forest productivity, herbivory, disturbance. Allen et al. suggest that authorities should focus on excluding ungulates on these highly productive regenerating forests rather than old-growth forests. I am disturbed by this suggestion. It exposes the most biologically diverse forests to continuing damage.
Data gaps
New Zealand has many long-lived, slow-growing tree species. Recruitment of understory trees is already low across both main islands. This situation has been attributed to ungulate browsing. Over centuries, this might result in shifts in the canopy composition. Allen et al. call for additional research to increase our understanding of how browsing and other short-and long-term drivers affect the regeneration of large trees. Also, data on soil CO2 emissions needs better integration.
Australian brushtail possum; photo by Peter Firminger via Flickr
The study did not consider the impact of other introduced mammals, such as feral pigs (Sus scrofa), rodents, and Australian brushtail possum (Trichosurus vulpecula). The possum is known to damage New Zealand trees. The scientists did not explain this omission; I assume it might have been the result of either lack of resources to support a broader study or differences in management strategies – or both?
I note that the study also did not address the extent to which non-native pathogens threaten these large trees. In response to my query, Kara Allen said that their plots did not include many kauri (Agathis australis) trees, so the severe dieback disease caused by Phytophthora agathidicida did not affect their results. Naturally regenerating kauri is limited to a small area of warm temperate rainforests located at the top of the North Island. So kauri potentially play a relatively small role in terms of overall carbon stocks in New Zealand’s forests. On the other hand, Allen says thatmyrtle rust (Austropuccinia psidii) could have a major impact on New Zealand forests’ carbon storage. Trees in the host family, Myrtaceae, are ecologically important across both islands. Also, they comprise a large portion of overall forest carbon stocks (ranked in the top 5 largest families for above- and belowground biomass). An example is southern rata (Mterosideros umbellata), which are preferentially fed on by Australian brush possum.
southern rata; photo by Stan Shebs
Bernd Blossey, (free access!) who has long studied the role of high deer populations in North American forests, praises the study’s attempt to measure data, not just rely on models, and its inclusion of soil. However, he notes other limitations of the New Zealand study:
The small exclosures (20 x 20 m) are subject to edge effects. Some of Blossey’s exclosures occupy 2 hectares.
Twenty years is too short a time for analysis of such long-term processes as carbon sequestration and regeneration of slow-growing trees. Therefore, any results must be considered preliminary. Furthermore, no one recorded any differences in carbon sequestration of the paired plots at the time the exclosures were set up.
There’s no mention of possible impacts by introduced earthworms.
Dr. Blossey recognizes that the current study’s authors cannot re-do actions taken decades in the past. Still, the data gaps reduce the value of the findings.
I conclude that uncertainties continue due to: the long timelines of species’ regeneration and growth to full sizes; the requirement for large exclosures; the complexity of factors affecting carbon sequestration; and probably other influences.. Managers trying to maximize carbon sequestration are forced to act without truly knowing the best strategy or how their actions will affect the future.
For more about invasive mammals’ impacts in U.S. forests, also see the study by USFS scientists, Poland et al. (full citation listed in sources). One can enter “mammal” in the search box for the on-line PDF.
SOURCES
Allen, K., P.J. Bellingham, S.J. Richardson, R.B. Allen, L.E. Burrows, F.E. Carswell, S.W.Husheer, M.G. St. John, D.A. Peltzer, M. Whenua. 2023. Long-term exclusion of invasive ungulates alters tree recruitment and functional traits but not total forest carbon. Ecological Applications. 2023; e2836. https://onlinelibrary.wiley.com/r/eap
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. The on-line version as at https://link.springer.com/book/10.1007/978-3-030-45367-1
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
black locust – one of the most widespread invasive tree species on Earth; photo via Flickr
In recent years there has been an encouraging effort to examine bioinvasions writ large see earlier blogs re: costs of invasive species – here and here. One of these products is the Routledge Handbook of Biosecurity and Invasive Species (full citation at end of this blog). I have seen only the chapter on bioinvasion in forest ecosystems written by Sitzia et al. While they describe this situation around the globe, their examples are mostly from Europe.
Similar to other overviews, this article re-states the widely-accepted attribution of rising numbers of species introductions to globalization, especially trade. In so doing, Sitzia et al. assert that the solution is not to curtail trade and movement of people, but to improve scientific knowledge with the goal of strengthening biosecurity and control programs. As readers of this blog know, I have long advocated more aggressive application of stronger restrictions on the most high-risk pathways. Still, I applaud efforts to apply science to risk assessment.
Sitzia et al. attempt to provide a global perspective. They remind readers that all major forest ecosystems of Earth are undergoing significant change as a result of conversion to different land-uses; invasion by a wide range of non-native introduced species—including plants, insects, and mammals; and climate change. These change agents act individually and synergistically. Sitzia et al. give greater emphasis than other writers to managing the tree component of forests. They explain this focus by asserting that forest management could be either the major disturbance favoring spread of non-native species or, conversely, the only way to prevent further invasions. They explore these relationships with the goal of improving conservation of forest habitats.
Japanese stiltgrass invasion; photo by mightyjoepye via Flickr
Sitzia et al. focus first on plant invasions. They contend that – contrary to some expectations – plants can invade even dense forests despite competition for resources. They cite a recent assessment by Rejmánek & Richardson that identified 434 tree species that are invasive around Earth. Many of these species are from Asia, South America, Europe, and Australia. These non-native trees can drive not only changes in composition but also in conservation trajectories in natural forests. However, the example they cite, Japanese stilt grass (Microstegium vimineum) in the United States, is not a tree! Sitzia et al. note that in other cases it is difficult to separate the impacts of management decisions, native competitive species, and non-native species.
Sitzia et al. note that plant invasions might have a wide array of ecological impacts on forests. They attempt to distinguish between
“drivers” of environmental change – including those with such powerful effects that they call them “transformers”;
“passengers” whose invasions are facilitated by other changes in ecosystem properties; and
“backseat drivers” that benefit from changes to ecosystem processes or properties and cause additional changes to native plant communities.
An example of the last is black locust (Robinia pseudoacacia). This North American tree has naturalized on all continents. It is a good example of the management complexities raised by conflicting views of an invasive species’ value, since it is used for timber, firewood, and honey production.
Sitzia et al. then consider invasions by plant pathogens. They say that these invasions are one of the main causes of decline or extirpations in tree populations. I applaud their explicit recognition that even when a host is not driven to extinction, the strong and sudden reduction in tree numbers produces significant changes in the impacted ecosystems.
American chestnut – not extinct but ecological role gone; photo by F.T. Campbell
Sitzia et al. contend that social and economic factors determine the likelihood of a species’ transportation and introduction. Specifically, global trade in plants for planting is widely recognized as being responsible for the majority of introductions. Introductions via this pathway are difficult to regulate because of the economic importance (and political clout) of the ornamental plants industry, large volumes of plants traded, rapid changes in varieties available, and multiple origins of trade. As noted above, the authors seek to resolve these challenges by improving the scientific knowledge guiding biosecurity and control programs. In the case of plant pathogens, they suggest adopting innovative molecular techniques to improve interception efficiency, esp. in the case of latent fungi in asymptomatic plants.
The likelihood that a pathogen transported to a new region will establish is determined by biogeographic and ecological factors. Like other recent studies, Sitzia et al. attempt to identify important factors. They name a large and confusing combination of pathogen- and host-specific traits and ecosystem conditions. These include the fungus’ virulence, host specificity, and modes of action, reproduction, and dispersal, as well as the host’s abundance, demography, and phytosociology. A key attribute is the non-native fungus’ ability to exploit micro-organism-insect interactions in the introduced range. (A separate study by Raffa et al. listed Dutch elm disease as an example of this phenomenon.) I find it interesting that they also say that pathogens that attack both ornamental and forest trees spread faster. They do not discuss why this might be so. I suggest a possible explanation: the ornamental hosts are probably shipped over wide areas by the plant trade.
surviving elms in an urban environment; photo by F.T. Campbell
Sitzia et al. devote considerable attention to bioinvasions that involve symbiotic relationships between bark and ambrosia beetles and their associated fungi. These beetles are highly invasive and present high ecological risk in forest ecosystems. Since ambrosia beetle larvae feed on symbiotic fungi carried on and farmed by the adults inside the host trees, they are often polyphagous. Bark beetles feed on the tree host’s tissues directly, so they tend to develop in a more restricted number of hosts. Both can be transported in almost all kinds of wood products, where they are protected from environmental extremes and detection by inspectors. Sitzia et al. specify the usual suspects: wood packaging and plants for planting, as ideal pathways. These invasions threaten indigenous species by shifting the distribution and abundance of certain plants, altering habitats, and changing food supplies. The resulting damage to native forests induces severe alterations of the landscape and causes economic losses in tree plantations and managed forests. The latter losses are primarily in the high costs of eradication efforts – and their frequent failure.
Eucalyptus plantation in Kwa-Zulu-Natal, South Africa; photo by Kwa-Zulu-Natal Department of Transportation
Perhaps their greatest contribution is their warning about probable damage caused by invasive forest pests in tropical forests. (See an earlier blog about invasive pests in Africa.) Sitzia et al. believe that bark and ambrosia beetles introduced to tropical forests threaten to cause damage of the same magnitude as climate change and clear cutting, but there is little information about such introductions. Tropical forests are exposed to invading beetles in several ways:
1) A long history of plant movement has occurred between tropical regions. Sitzia et al. contend that the same traits sought for commercial production contribute to risk of invasion.
2) Logging and conversion of tropical forests into plantation forestry and agriculture entails movement of potentially invasive plants to new areas. Canopies, understory plant communities, and soils are all disturbed. Seeds, insects, and pathogens can be introduced via contaminated equipment.
3) Less developed nations are often at a disadvantage in managing potential invasion. Resources may be fewer, competing priorities more compelling, or potential threats less obvious.
Sitzia et al. call for development of invasive species management strategies that are relevant to and realistic for less developed countries. These strategies must account for interactions between non-native species and other aspects of global environmental change. Professional foresters have a role here. One clear need is to set out practices for dealing with conflicts between actors driven by contrasting forestry and conservation interests. These approaches should incorporate the goals of shielding protected areas, habitat types and species from bioinvasion risk. Sitzia et al. also discuss how to address the fact that many widely used forestry trees are invasive. (See my earlier blog about pines planted in New Zealand.)
planted forest in Sardinia, Italy; photo by Torvlag via Flickr
In Europe, bark beetle invasions have damaged an estimated ~124 M m2 between 1958 and 2001. Sitzia et al. report that the introduction rate of non-native scolytins has increased sharply. As in the US, many are from Asia. They expect this trend to increase in the future, following rising global trade and climate change. Southern – Mediterranean – Europe is especially vulnerable. The region has great habitat diversity; a large number of potential host trees; and the climate is dry and warm with mild winters. The region has a legacy of widespread planting of non-native trees which are now important components of the region’s economy, history and culture. These include a significant number of tree species that are controversial because they are – or appear to be – invasive. Thus, new problems related to invasive plants are likely to emerge.
Noting that different species and invasion stages require different action, Sitzia et al. point to forest planning as an important tool. Again the discussion centers on Europe. Individual states set forest policies. Two complications are the facts that nearly half of European forests are privately owned; and stakeholders differ in their understanding of the concept of “sustainability”. Does it mean ‘sustainable yield’ of timber? Or providing multiple goods and services? Or sustaining evolution of forest ecosystems with restrictions on the use of non-native species? Resolving these issues requires engagement of all the stakeholders.
Sitzia et al. say there has recently been progress. The Council of Europe issued a voluntary Code of Conduct on Invasive Alien Trees in 2017 that provides guidelines on key pathways. A workshop in 2019 elaborated global guidelines for the sustainable use of non-native tree species, based on the Bern Convention Code of Conduct on Invasive Alien Trees. The workshop issued eight recommendations:
Use native trees, or non-invasive non-native trees;
Comply with international, national, and regional regulations concerning non-native trees;
Be aware of the risk of bioinvasion and consider global change trends;
Design and adopt tailored practices for plantation site selection and silvicultural management;
Promote and implement early detection and rapid response programs;
Design and adopt practices for invasive non-native tree control, habitat restoration, and for dealing with highly modified ecosystems;
Engage with stakeholders on the risks posed by invasive NIS trees, the impacts caused, and the options for management; and
Develop and support global networks, collaborative research, and information sharing on native and non-native trees.
SOURCE
Sitzia, T., T. Campagnaro, G. Brundu, M. Faccoli, A. Santini and B.L. Webber. 2021 Forest Ecosystems. in Barker, K. and R.A. Francis. Routledge Handbook of Biosecurity and Invasive Species. ISBN 9780367763213
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
spotted lanternfly – could we have predicted its arrival? Its Impacts? Photo by Holly Ragusa, Pennsylvania Department of Agriculture
As readers of my blogs know, I wish to prevent introduction and spread of tree-killing insects and pathogens and advocate tighter and more pro-active regulation as the most promising approach. I cannot claim to have had great success.
Of course, international trade agreements have powerful defenders and the benefit of inertia. And in any case, prevention will be enhanced by improving the accuracy of predictions as to which specific pests are likely to cause significant damage, which are likely to have little impact in a naïve ecosystem. This knowledge would allow countries to can then focus their prevention, containment, and eradication efforts on this smaller number of organisms.
I applaud a group of eminent forest entomologists and pathologists’ recent analysis of widely-used predictive methods’ efficacy [see Raffa et al.; full citation at the end of this blog]. I am particularly glad that they have included pathogens, not just insects. See earlier blogs here, here,here, and here.
I review their findings in some detail in order to demonstrate their importance. National and international phytosanitary agencies need to incorporate this information and adopt new strategies to carry out their duty to protect Earth’s forests from devastation by introduced pests.
Raffa et al. note the usual challenges to plant health officials:
the high volumes of international trade that can transport tree-killing pests;
the high diversity of possible pest taxa, exacerbated by the lack of knowledge about many of them, especially pathogens;
the restrictions on precautionary approaches imposed by the World Trade Organization’s Sanitary and Phytosanitary Agreement (the international phytosanitary system) – here, here, and here.
the high cost and frequent failure of control efforts.
ash trees killed by emerald ash borer; Mattawoman Creek, Maryland; photo by Leslie A. Brice
The Four Approaches to Predicting Damaging Invaders
At present, four approaches are widely used to predict behavior of a species introduced to a naïve environment:
(1) pest status of the organism in its native or previously invaded regions;
(2) statistical patterns of traits and gene sequences associated with high-impact pests;
(3) sentinel plantings to expose trees to novel pests; and
(4) laboratory tests of detached plant parts or seedlings under controlled conditions.
Raffa et al. first identify each method’s underlying assumptions, then discuss the strengths and weaknesses of each approach for addressing three categories of biological factors that they believe explain why some organisms that are relatively benign, sparse, or unknown in their native region become highly damaging in naïve regions:
(1) the lack of effective natural enemies in the new region compared with the community of predators, parasites, pathogens, and competitors in the historical region (i.e., the loss of top-down control);
(2) the lack of evolutionary adaptation by naïve trees in the new region compared with long-term native interactions that select for effective defenses or tolerance (i.e., the loss of bottom-up control); and
(3) novel insect–microbe associations formed in invaded regions in which one or both members of the complex are non-native, resulting in increased vectoring of or infection courts for disease-causing pathogens (i.e., novel symbioses). I summarize these findings in some detail later in this blog.
Most important, Raffa et al. say none of these four predictive approaches can, by itself, provide a sufficiently high level of combined precision and generality to be useful in predictions. Therefore, Raffa et al. outline a framework for applying the strengths of the several approaches (see Figure 4). The framework can also be updated to address the challenges posed by global climate change.
Raffa et al. repeatedly note that lack of information about pests undercuts evaluation efforts. This is especially true for pathogens and the processes determine which microbes that are innocuous symbionts in co-evolved hosts become damaging pathogens when introduced to naïve hosts in new ecosystems.
Findings in Brief
Raffa et al. found that:
Previous pest history in invaded environments provides greater predictive power than population dynamics in the organism’s native regions.
Models comparing pest–host interactions across taxa are more predictive when they incorporate phylogenies of both pest and host. Traits better predict a pest’s likelihood of transport and establishment than its impact.
Sentinel plantings are most applicable for pests that are not primarily limited to older trees. Ex patria sentinel plantations are more likely to detect pest species liberated by loss of bottom-up controls than top-down controls, i.e., most fungi and woodborers but not insect defoliators.
Laboratory tests are most promising for pest species whose performance on seedlings and detached parts (e.g., leaves) accurately reflects their performance on live mature trees. They are thus better at predicting impacts of insect folivores and sap feeders than woodborers or vascular wilt pathogens.
Raffa et al. also ask some fundamental questions:
How realistic is it to expect reliable predictions, given the uniqueness of each biotic system?
When should negative data – lack of data showing a species is invasive – justify decisions not to act? Especially when there are so many data gaps?
Who should make decisions about whether to act? How should the varying values of different social sectors be incorporated into decisions?
Raffa et al. identify critical areas for improved understanding:
1) Statistical tools and estimates of sample size needed for reliable forecasts by the various approaches.
2) Reliability, breadth, and efficiency of bioassays.
3) Processes by which some microorganisms transition from saprophytic to pathogenic lifestyles.
4) Procedures for scaling up results from bioassays and plantings to ecosystem- and landscape-level dynamics.
5) Targetting and synergizing predictive approaches and methods for more rapid and complete information transfer across jurisdictional boundaries.
I am struck by two generalizations:
While most introduced forest insects are first detected in urban areas, introduced pathogens are more commonly detected in forests. I suggest that more intensive surveys of urban trees and “sentinel gardens” might result in detection of pathogens before they reach the forest.
Enemy release is rarely documented as the primary basis for pathogens that cause little or no impact in their native region but become damaging in an introduced region. Enemy release appears generally more important with folivores and sap feeders than with woodborers.
Detailed Evaluation of Predictive Methodologies
white pine blister rust-killed whitebark pine at Crater Lake National Park; photo by F.T. Campbell
Empirical assessment of pest status in previously occupied habitats
This is the most commonly applied method now, partly because it seems to follow logically from the World Trade Organization’s requirement that national governments provide scientific evidence of risk to justify adopting phytosanitary measures. The underlying assumption is that species that have caused damage in either their native or previously invaded ranges are those most likely to cause damage if introduced elsewhere. The corollary is that species that have not previously caused damage are unlikely to cause significant harm in a new ecosystem.
As noted above, Raffa et al. found that a species’ damaging activity in a previously invaded area can help indicate likely pest status in other regions. However, its status — pest or not — in its native range is not predictive. See Table 1 for numerous examples of both pests and non-pests. For example, Lymantria dispar has proved damaging in both native and introduced ranges. Ips typographus has not invaded new territories despite being damaging in its nature range and frequently being transported in wood. White pine blister rust is not an important mortality source on native species in its native range but is extremely damaging in North America.
Raffa et al. also note the importance of whether effective detection and management strategies exist in determining a pest’s impact ranking. Insects are more easily detected than pathogens; some respond to long distance attractants such as pheromones or plant volatiles. These methods can include insect vectors of damaging pathogens.
Re: the difficulty of assessing insect–microbe associations, they name several examples of symbionts which have caused widespread damage to naïve hosts: laurel wilt in North America; Sirex noctilio and Amylosterum areolatum around the Southern Hemisphere; Monochamus spp. and Bursaphelenchus xylophilus in Asian and European pines. Dutch elm disease illustrates a widespread epidemic caused by replacement of a nonaggressive native microorganism in an existing association with a non-native pathogen. Beech bark diseaseresulted from independent co-occurrence of an otherwise harmless fungus and harmless insect.
In sum, “watch” lists are disappointingly poor at identifying species that are largely benign in their native region but become pests when transported to naive ecosystems. Many of our most damaging pests are in this group. Raffa et al. note that this is not surprising because naïve systems lack the very powerful top-down, bottom-up, and lateral forces that suppress pests’ populations in co-adapted system. Countries often try to overcome this uncertainty by shifting to pathway mitigation and other “horizontal measures” – as I have often advocated. Raffa et al. emphasize that such approaches are costly to implement and constrain free trade.
Predictive models based on traits of pests and hosts
Predictive models provide the most all-encompassing and logistically adaptable of the forecasting approaches. Typically, models consider various components of risk, e.g. probability of transport, probability of establishment, anticipated level of damage.
The overriding assumption is that patterns emerging from either previous invasions or basic biological relationships can provide reliable predictions of impacts that might result from future invasions. However, Raffa et al. note that the models’ reliability and specificity are hampered by small sample sizes and data gaps.
They found that specific life history traits have proved to be more predictive of insect — and to a lesser extent fungal – establishment than of impact. Earlier studies [Mech et al. (2019) and Schulz et al. (2021)] found no association between life history traits and impacts for either conifer-feeding or angiosperm-feeding insects.
Some traits of pathogens have been linked to invasion success, e.g., dispersal distance, type of reproduction, spore characteristics, and some temperature characteristics for growth and parasitic specialization. Raffa et al. say that root-infecting oomycete pathogens have a broader host range and invasive range than those that attack aboveground parts. Oomycetes that grow faster and produce thick-walled resting structures have broader host ranges. Phenotypic plasticity is also important. Raffa et al. say that those organisms that require alternate hosts can be limited in their ability to establish. However, they don’t mention that – once introduced — they can have huge impacts, as the example of white pine blister rust illustrates.
Raffa et al. say that phylogenetic distance of native and introduced hosts is more predictive for foliar ascomycetes than for basidiomycete and oomycete pathogens with broad host ranges. They suggest predictive ability can be improved by incorporating other factors, e.g., feeding guild. They note that the findings of Mech et al. and Schulz et al. (see links above) show the importance ofboth host associations with pests and phylogenetic relationships between native and naïve hosts for predicting impacts.
Geography is important: while there is a greater chance of Northern Hemisphere pests invading in the same hemisphere, this is not universal, as shown by Sirex (of course, the woodwasp is attacking hosts native to the Northern Hemisphere – pines).
Genomic analyses have been used more often with pathogens. There are two general approaches:
trees killed by chestnut blight; USDA Forest Service photo
1) Comparing the genomes of different species to identify the determinants associated with certain traits or lifestyles. For example, a post hoc analysis of the genus Cryphonectria could distinguish nonpathogenic species from the chestnut blight fungus C. parasitica.
2) Using genomic variation within a single species to identify markers associated with traits. Genome sequencing of a worldwide collection of the pathogens that cause Dutch elm disease revealed that some genome regions that originated from hybridization between fungal species contained genes involved in host–pathogen interactions and reproduction, such as enhanced pathogenicity and growth rate.
Raffa et al. point out that the growth of databases will facilitate genomic approaches to identify important invasiveness and impact traits, such as sporulation, sexual reproduction, and host specificity.
At present, Raffa et al. believe that models based on traits, phylogeny, and genomics offer potential for a rapid first pass to predicting levels of pest damage. However, assessors must first have a list of candidate pest species and detailed information about each. Plus there is still too much uncertainty to rely exclusively on the models.
Sentinel trees
Raffa et al. say that sentinel trees can potentially provide the most direct tests of tree susceptibility and the putative impact of introduced pests. Three types of plantations offer different types of information:
In patria sentinels [= sentinel nurseries] = native trees strategically located in an exporting country and exposed to native pests. The intention is to detect problematic hitchhikers before they are transported to a new region. These plantings are useful for commodity risk assessment. However, all the taxa associated with the sentinel trees must be identified to ascertain whether they can become a threat to plants in the new ecosystem.
Ex patria plantings [= sentinel plantations] = trees from an importing country are planted in an exporting country with the aim of assessing new pest–host associations. These plantings are most useful for identifying threats that arise primarily from lack of coevolved host tree resistance (i.e., loss of bottom-up control). They cannot predict the effects of lack of co-adapted natural enemies in the importing region (i.e., loss of top-down control). Plantings are thus more helpful in predicting impacts by pathogens and woodborers than folivores and sap feeders. However, ex patria plantings cannot predict pest problems that arise from novel microbial associations, or increased susceptibility to native pests.
Trees in botanic gardens, arboreta, large-scale plantations, and urban parks and yards can provide information on both existing native-to-native associations and new pest–host associations. Analyzing these plantings can be useful for studying host-shift events and novel pest–host associations. Again, all the taxa associated with the sentinel trees must be identified to ascertain whether they can become a threat to plants in the new ecosystem. Monitoring these planting have detected previously unknown plant–host associations (such as polyphagous shot hole borer and tree species in California and South Africa), and entirely unknown taxa. Pest surveillance in urban areas can also facilitate early detection, thereby strengthening the possibility of eradication.
PSHB attack on Erythrina caffri; photo by Paap
Sentinel tree programs are limited by 1) small sample sizes; 2) immature trees; and 3) the fact that trees planted outside their native range might not be accurate surrogates for the same species in native conditions. Some of these issues can be reduced by establishing reciprocal international agreements among trading partners; the International Plant Sentinel Network helps to coordinate these collaborations.
Botanic gardens and arboreta have the advantage of containing adult trees; this is important because pest impacts can vary between sapling to mature trees. However, they probably contain only a few individuals per plant species, usually composed of narrow genetic base.
Large-scale plantations of exotic tree species, e.g., exotic commercial plantations, comprise large numbers of trees planted over large areas with varied environmental conditions, and they stand for longer times. Still, they commonly have a narrow genetic base that might not be representative of wild native plants. Also, only a few species are represented in commercial plantations.
Raffa et al. report that experience in commercial Eucalyptus plantations in Brazil alerted Australia to the threat from myrtle rust (Austropuccinia psidii). However, in an earlier blog I showed that Australia did not act quickly based on this knowledge.
Laboratory assays using plant parts or seedlings
Laboratory tests artificially challenge seedlings, plant parts (e.g., leaves, branches, logs), or other forms of germplasm of potential hosts to determine their vulnerability. These tests are potentially powerful because they are amenable to experimental control, standardized challenge, and replication. They also avoid many of the logistical constraints of sentinel plantings. Finally, they can be performed relatively rapidly.
The key underlying assumption is that results can be extrapolated to predict injury to live, mature treesunder natural conditions. The validity of this assumption depends on the degree to which exogenous biotic and abiotic stressors affect the outcomes. Raffa et al. report that environmental stressors tend to more strongly influence tree interactions with woodborers than folivores.
These assumption are more likely to be met by pathogens that infect shoots or young tissues, such as the myrtle rust pathogenAustropuccinia psidii, ash dieback pathogen Hymenoscyphus fraxineus, and the sudden oak death pathogen Phytophthora ramorum.
The host range of and relative susceptibilities to insects is usually tested on twigs bearing foliage for defoliators and sap suckers; bark disks, logs, or branches for bark beetles, ambrosia beetles, and wood borers. These methods do not work as well for bark beetle species that attack mature trees in which active induced responses and transport of resins through established ducts are critically important.
The major advantages of laboratory tests is that they readily incorporate both positive (known hosts) and negative (known nonhosts) controls, can provide a range of environmental conditions, can be performed relatively rapidly, are statistically replicable at relatively low costs, and can test multiple host species and genotypes simultaneously. The ability to statistically replicate a multiplicity of environmental combinations and species is particularly valuable for evaluating relationships under anticipated future climatic conditions.
However, there are several important limitations. In testing pathogens, environmental conditions required for infection are often unknown. Choice of non-conducive conditions might result in false negatives; choice of too-conducive conditions might result in exaggerating the likelihood of infection. Results of tests of insect pests can vary depending on whether the insects are allowed to choose among potential host plants. Other complications arise when the pest being evaluated requires alternate hosts. In addition, seedlings are not always good surrogates for mature trees – especially as regards pathogens and bark, wood-boring and root collar insects. Folivores are less affected by conditions. Plus, the costs can be significant since they involve maintaining a relatively large number of viable and virulent pathogen cultures, insects, and candidate trees in quarantine.
Finally, although lab assays are well suited for identifying new host associations, results might not be amenable to scaling up to predict a pest’s population-level performance in a new ecosystem. Scaling up is especially problematic for those insect species whose dynamics are strongly affected by trophic interactions.
SOURCE
Raffa, K.F., E.G. Brockerhoff, J-C Gregoire, R.C. Hamelin, A.M. Liebhold, A. Santini, R.C. Venette, and M.J. Wingfield. 2023. Approaches to Forecasting Damage by Invasive Forest P&P: A Cross-Assessment. BioScience Vol. 73 No. 2: 85–111 https://doi.org/10.1093/biosci/biac108
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
sooty bark disease; photo by Vincet Gaucet via Bugwood
Recently, plant pathologists have paid more attention to pathogens — native to eastern North America — now killing trees on the West Coast. Years ago, the likelihood of such “domestic invaders” was hotly debated. However, these new detections provide examples of native micro-organisms that are apparently benign in the ecosystems in which they evolved, but that cause disease when moved to relatively nearby — but naïve — environments. I would argue that it is the absence of co-evolution of pathogen and host, not distance, that matters. In both cases, environmental stress on the trees appears to play a significant role. Such stress is expected to increase as the climate changes.
[In the past, these issues arose re: goldspotted oak borer – introduced to California from Arizona; and thousand cankers disease – introduced from Arizona to Western states and now to the East.]
Little is known about these diseases. So far, scientists have examined the pathogens’ impacts more thoroughly on plantings that are introduced to the location, so already outside of the forest biomes in which they evolved. Less attention has been paid to hosts native to the regions newly affected. I find this disturbing because I am most concerned about the possible impact to native ecosystems.
I will describe two such diseases. Both attack native tree species, not just the exotic trees described in research. Both were first detected in the Pacific Coast states in the late 1960s – i.e., more than 50 years ago. Why are they becoming prominent now – is it changes in the climate? Evolution? Just slow adaptation?
Example One – Sooty Bark Disease
Sooty-bark disease (Cryptostroma corticale) of maples (Acer spp.) is native to the Great Lakes region, where it causes no problems. However, it has been introduced to the West Coast, where all sources agree that the disease kills trees stressed by heat and drought. These stresses are expected to increase as the climate changes. The disease is also a serious threat to human health. The fungus’ spores can cause serious pulmonary disease in humans.
Sooty bark disease was detected in Pullman, Washington, near the Idaho border, in 1968. Now it is more widespread: it was detected in the Seattle area as of 2020 and the Sacramento area of California in 2019 [Curtis Ewing pers. comm.]. Sooty-bark disease has also spread to at least ten countries in Europe, ranging from the United Kingdom to Italy and Bulgaria.
Sources in Washington say hosts include several non-native species widely planted in the area – sycamore maples (Acer pseudoplatanus) [Chastagner], Norway maples (Acer platanoides), Japanese maple (A. palmatum), and horsechestnut (Aesculus hippocastanum) [Chastagner and Washington State University]. More troubling is the fact that several native tree species are also hosts. Big leaf maple (A. macrophyllum), and Pacific dogwood (Cornus nuttallii) are among the most significant. [Big leaf maple is a large hardwood tree in a region dominated by conifers. Pacific dogwood has already been decimated by the introduced disease dogwood anthracnose.] Sources in California add two other maples, these native to eastern North America, red (A. rubrum) and silver (A. saccharinum) maples.
bigleaf maples in Olympic National Park; NPS photo
The fungus initiates infection in a tree’s small branches, then spreads into the heartwood and both up and down the tree. It might also invade pruning wounds. The fungus grows more rapidly at higher temperatures. It is also facilitated by drought stress. When the fungus grows out to the bark, it causes the bark to blister; it then forms spore-forming structures. The spores are dispersed by wind. (Chastagner)
Washington State University’s Ornamental Plant Pathology division has called for more research on all aspects of the disease in trees:
Distribution and spread of the disease;
Plant species susceptibility and host range;
Diagnostic methods and molecular approaches to improve diagnostic efficiency and capacity;
Pathogen life history and genetic diversity;
Factors that affect disease development and vulnerability (site, stress, age of host, etc.);
Potential of human mediated dispersal and vectors such as pruning tools;
Best management practices and worker protection.
So far, little has been done. Scientists in the Pacific Northwest have received a small amount of funding from the USDA Forest Service Forest Health Protection Emerging Pests program link + blog on appropriations to increase diagnostic services and to surveys trees elsewhere in the Puget Sound region.
Example Two – Bacterial Pathogen on Oaks
drippy blight disease on northern red oak; photo by Rachel Sitz, USDA Forest Service; via Bugwood
There is always concern about threats to oaks (Genus Quercus) because of their ecological, economic, and social importance. As Kozhar et al. point out, this genus is one of the most important groups of trees in many regions of the Northern Hemisphere. In North America specifically, oak forests compose a significant part of many forest ecosystems, especially in the East. California has 20 native species, Colorado has one. In addition, oaks are also often planted as shade trees in urban environments, which has resulted in movement of oak species to new geographic areas. There where they experience different environmental conditions, they might find new pests or alert us to the effects of climate change.
The bacterial oak pathogen Lonsdalea quercina is indigenous to the native range of northern red oak (Quercus rubra) in eastern North America. There, it does not cause disease in its co-evolved host. However, it has recently caused two outbreaks in the West – in California and Colorado. In the latter, trees themselves can die; in the former, acorns are damaged, threatening forest regeneration. Other Lonsdalea species have caused similar tree diseases in Europe.
California
In California, the bacterium has been present since at least 1967. It infects acorns of native oaks, including coast live oak (Q. agrifolia), Q. parvula (presumably the mainland subspecies, also named Q. p. var. shrevei),and interior live oak (Q. wislizeni). The Morton Arboretum link says Q. parvula (presumably – again – the mainland subspecies) is currently threatened by sudden oak death (SOD). Coast live oak has also been highly affected by sudden oak death (SOD), DMF but it is not considered to be threatened. I expect – but sources don’t say – that the bacterium is affecting these species’ reproduction.
Various genotypes of L. quercina are randomly distributed across trees in both native and human-altered habitats and among all host species. Kohzar et al. say this is not surprising since all the host oak species are native to the region. Coast live oak is a major component of native forests and is also widely planted as a shade tree in residential areas. Furthermore, there has been no attempt to restrict the pathogen’s movement by adopting quarantines or other measures by phytosanitary agencies.
As a result, the inoculum can be moved across large distances by insects, birds, small mammals, and humans.
Bacterial pathogens can be associated with insects, relying on their feeding sites and other wounds to facilitate entry to the host’s tissues or for dissemination among hosts. In California, L. quercina might enter host tissue via wounds made by acorn weevils, filbertworms, and some cynipid wasps Kohzar et al.).
Colorado
The situation in Colorado is different. Significant dieback of exotic oaks planted in the state came to attention in the early 2000s. The hosts include non-native northern red oak, pin oak (Q. palustris), and Shumard oak (Q. shumardii). In Colorado, the bacterium causes “drippy blight disease” on the trees, not the acorns. The disease causes abundant ooze on symptomatic tissue. There has been a significant increase in tree mortality – with associated removal costs. The bacterium also has been found attacking Colorado’s one native oak, Q. gambelii; in this case, the pathogen attacks the acorns rather than the tree.
Due to small sample sizes, Kohzar et al. were unable to answer three key questions:
whether the Colorado L. quercina population comprises a new taxonomic species;
whether genetic variation in the bacterial populations are explained by the habitat (native or human-altered) or host; and
whether the L. quercina infections on native Q. gambelii serves as an inoculum reservoir for planted Q. rubra hosts or vice versa.
gambel oak; photo by Dave Powell, USDA Forest Service (retired); via Bugwood
Surprisingly, despite its more recent emergence, the Colorado population of L. quercina has higher genetic diversity. Kohzar et al. suggest this might be due to repeated introductions of the bacterium on nursery stock brought in from the northern red oak’s native range in the East. [see below]
As in California, L. quercina infections are associated with insects, especially kermes scale (Allokermes galliformis). This insect does not travel long distances, which might help explain why the Colorado genotypes are limited to nearby trees, not dispersed randomly as in California.
However, kermes scale has been present in the state for far longer than the disease. The scale’s population spiked at the same time as the drippy blight outbreak was detected. Kohzar et al. could not determine whether the rise in scale populations and associated increase in number of entry points through feeding sites led to the increase of bacterial populations, or vice versa.
Kohzar et al. did determine that the Colorado populations of L. quercina were not introduced from California. They cannot explain the original introduction but think there might be continuing introductions from the native range of both northern red oak and L. quercina – the northeastern United States. They call for further studies to understand evolutionary relationships among L. quercina populations from different areas, including the native habitat of red oak in the East to clarify possible causes and sources of the recent outbreak of drippy blight in Colorado.
Role of Environmental Conditions
Kohzar et al. stress the importance of factors other than species’ introductions to new environments as the cause of emerging forest diseases. They say such other factors as changes in environmental conditions, new host-vector associations, cryptic disease agents (e.g., pathogens with a very long latency period or endophytes changing their behavior to pathogenic), hypervirulent strains of known pathogenic species, and/or newly emerging species of unknown origin as key factors leading to disease emergence in forest ecosystems around the globe.
In the case of L. quercina in California and especially Colorado, Kohzar et al. point to stress on the trees caused by new environmental factors, e.g., rapid climate change. [Of course, oaks from humid regions of eastern North America are already outside their natural habitat in much-dryer Colorado.] They support this conclusion by noting the simultaneous appearance of four new diseases caused by Lonsdalea species in different parts of the world during the 1990s and early 2000s. These were Lonsdalea quercina in Colorado; L.britannica on oaks in Great Britain; L. iberica in Spain; and L. populi bark canker on poplar species in Hungary, China, and Spain. All cause similar symptoms of drippy blight disease.
Ewing, Curtis. Entomologist, CalFire. Pers. comm. March 2023.
Kozhar, O., R.A. Sitz, R. Woyda, L. Legg, J.R. Ibarra Caballero, I.S. Pearse, Z. Abdo, J.E. Stewart. 2023. Population genomic analysis of an emerging pathogen Lonsdalea quercina affecting various species of oaks in western North America. BioRxiv https://www.biorxiv.org/content/10.1101/2023.01.20.524998v1
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
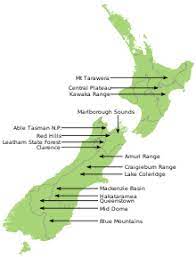
spread of non-native conifers in mountains of New Zealand; photos by Richard Bowman; New Zealand government website
As those of us who want to “do something” to counter bioinvasions struggle to mobilize both the resources and the political will necessary, I rejoice that more studies are examining what factors affect “social license” [= public approva] for such programs. One such study was recently published in New Zealand — Mason et al. (full citation at the end of the blog). New Zealand enjoys a greater appreciation of the uniqueness of its biology and awareness of invasive species’ impacts than the United States. However, their findings might provide useful guidance in the US and elsewhere.
Mason et al. sought to understand motivations of, and constraints on, those local groups responsible for controlling the spread of non-native conifers into New Zealand’s remnant native ecosystems. Non-forest ecosystems across much of the country are at risk of rapidly transforming into exotic conifer forests. For these reasons, authorities are pressing for timely removal of existing seed sources, that is, mature non-native conifer trees of several species. The blog I posted earlier apparently describes effects of conifer invasions in lowland ecosystems, whereas the Programme described here is focused on high-elevation systems.
The eradication effort in the study is the National Wilding Conifer Control Programme, establishedin 2016. A large increase in funding provided during the COVID-19 lockdown made it practical to try to eradicate seed sources from large swathes of vulnerable land. The Programme coordinates control efforts across the country, working across property and land-tenure boundaries. Landowners are expected to cover 20% of the cost of removing conifers from their land. Since removing all seed sources of high-risk conifer species from the landscape is key to achieving long-term goals, success is unlikely if significant seed sources are allowed to persist.
Mason et al. combined workshops, questionnaires, and site visits to gather data on particular aspects of this Programme. They found that social resistance, rather than lack of scientific knowledge, was often the main barrier to success in managing widespread invasive species. The authors do not address whether the fact that only 30 people provided information for their study might undermine the reliability of their findings.
map of conifer wilding sites; adapted from Wilding conifers – New Zealand history and research background, a presentation by Nick Ledgard at the “Managing wilding conifers in New Zealand – present and future” workshop (2003)
The authors suggest that the main benefit of scientific information might be to increase stakeholders’ support for management interventions — rather than to guide manager’ decisions about which strategies to pursue. To support social license, invasive species research programs might need to focus not only on cost-effective control technologies and strategies, but – perhaps especially — the benefits (both tangible and intangible) of invasive species control for society.
Mason et al. found that people were motivated to combat conifer invasions by impacts with direct influence on humans or human activities (e.g., reduced water yield, damage to infrastructure from wildfires, reduced tourist activities due to landscape transformation) and also by impacts affect ecosystems (e.g., impacts on biodiversity, aquatic ecosystems and landscapes).
People objected to control or eradication programs primarily because of social concerns. These included the unwillingness of landowners to participate and regulatory frameworks that had perverse incentives.
Mason et al. called for greater efforts by scientists to persuade stakeholders[p1] to allow removal of “wilding” conifers from private land and development of more appropriate regulations. They found that forecasting models were particularly effective in persuading people to support these efforts. It seems to me that outreach teams might need “translators” to convert scientists’ findings to information that would be more useful by stakeholders.
The authors concede that the “wilding conifer” situation has unique attributes. First, invading conifers present a stark, easily seen difference between native and invaded ecosystems. Second, some – but not all—stakeholders appreciate the uniqueness of New Zealand’s biomes. Third, the impacts of conifer invasion are sufficiently well known that they can be described succinctly and accurately.
Do these unique attributes undercut the relevance of this study to North America? It is still true that ongoing support from local stakeholders (including landowners and community groups) influences the effectiveness or profitability of managing invasive species. .It is also true that groups’ varying values affect willingness to support the activities.
Mason et al. think through the issue of stakeholders’ conflicting perspectives on the value of particular invasive species and the values threatened by that invader. These can include ethical or safety concerns around management methods, particularly regarding toxins and genetic modification. Economoic costs are also a factor – especially if the landowner must pay all or some of them.
I find it interesting that the government simultaneously funded a 5-year research program to study various issues regarding the spread, ecosystem impacts, and control of wilding conifers. The result is the Mason et al. study discussed here. I wish the U.S. funded independent analyses of its invasive species programs!
Pinus contorta – the most rapidly growing Pinus introduced to New Zealand; photo by Walter Siegmund / Wikimedia
More Details, Policy Suggestions
Workshop attendees unanimously identified landscape impacts as a reason for controlling wilding conifers. This primarily concerned the loss of New Zealand’s visual heritage or cultural identity rather than loss of native species’ habitats. When the landowner was raised in Europe, these cultural or heritage values sometimes had the opposite effect, since they see conifer forests as important components of “natural” landscapes.
Currently, landowner funding and permission is required for conifer removal. Some individual landowners want to establish new forestry plantings. Some resist removal of existing forestry plantations (which provide income) and shelter belts (which provide shelter for livestock in high country landscapes). Some landowners were unwilling to pay their 20% of removal costs. Or they objected to certain conifer control methods—particularly helicopter spraying of herbicides. New Zealand’s regulatory process also requires years of negotiating to remove standing trees – further delaying any action. In theory, landowners who resist removal could be prosecuted under the Biosecurity Act. However, this approach has never been tried for removing wilding conifers.
Mason et al. suggested several changes in policy to overcome some of these barriers.
First, forestry consultants can “game” the wilding conifer “risk calculator” to obtain government approval to establish conifer plantations in high-risk environments. The authors suggest that authorities create a “liability calculator.” Under this system, landowners wishing to retain conifers on their land for whatever reason would be liable for any subsequent containment costs. However, developing such a tool requires more finely-scaled models of conifer spread.
Second, given the high costs of combatting invading conifers if seed sources are allowed to persist, they suggested that it might be more cost-effective for the control program to pay for plantation removal under New Zealand’s Emissions Trading Scheme.
Given the overwhelmingly social and regulatory nature of barriers to success, the primary role for scientific information is providing assessments of outcomes in the absence of wilding conifer control. Preferred messages were return-on-investment estimates and forecasts of ecosystem impacts, particularly relating to biodiversity loss, water yield reduction, and wildfire hazard. Forecasts were key to demonstrating that management interventions reduced future control costs and avoided environmental impacts which large sections of the community value (i.e. biodiversity loss, reduction in water yield and agricultural productivity, increased wildfire risks). Practitioners felt that forecasting models might also channel research toward areas of high uncertainty. Mason et al. recognize the difficulties presented by inherent complexity of ecological systems. However, they think “good practice” guidelines on forecasting are emerging.
The authors find that information content and presentation need to be tailored to the various audiences – most of whom lack experience in interpreting data from environmental forecasting models. They suggest that outreach materials focus on clear illustration of the tangible and intangible benefits of wilding conifer management rather than detailed explorations of scenarios. Participants suggested ways to improve the web tool to make it more accessible to a non-expert audience.
Mason et al. mention aspects that require balancing, but don’t suggest criteria for making these choices. They say it is important to include all relevant stakeholders in invasive species management governance bodies. The absence of stakeholders with positive attitudes to wilding conifer invasions led to unanticipated external social resistance to the Programme. They recognize that including stakeholders with conflicting interests might obstruct the decision-making process. Also, in areas where there has been success in containing conifers’ spread, people can’t see invading trees, so they don’t recognize the problem. They also note that existing data do not adequately recognize risks of spread from deliberately planted seed sources such as shelter-belts, plantations and amenity plantings. The authors do not discuss how to integrate these data into analyses and public outreach.
Finally, Mason et al. recognize that many other factors strongly influence stakeholders’ willingness to support invasive species control programs, especially the level of trust and strength of relationships between bioinvasion program staff and stakeholders.
Also, they suggest topics for future research: assessing how well forecasting models are integrated with communications with stakeholders; how qualitative and quantitative research methods in different fields might support one another; and empirical tests to measure the relative effects on social license of a) involving stakeholders in developing models, b) using forecasts to assess the consequences of different management decisions and, c) the usefulness of different methods for incorporating scientific information in stakeholder engagement.
SOURCE
Mason, N.W.H., Kirk, N.A., Price, R.J. et al. Science for social license to arrest an ecosystem-transforming invasion. Biol Invasions25, 873–888 (2023). https://doi.org/10.1007/s10530-022-02953-w
see also https://www.doc.govt.nz/nature/pests-and-threats/weeds/common-weeds/wilding-conifers/
Posted by Faith Campbell
What do YOU think about the role “social license” plays in US invasive species programs? We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm